Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2012; 3:285-291. doi:10.7150/jca.4537 This volume Cite
Research Paper
Analysis of Pancreatic Cancer Microenvironment: Role of Macrophage Infiltrates and Growth Factors Expression
Katarzyna Gardian ![]() , Sława Janczewska, Waldemar L. Olszewski, Marek Durlik
, Sława Janczewska, Waldemar L. Olszewski, Marek Durlik
Mossakowski Medical Research Centre Polish Academy of Sciences, Department of Surgical Research and Transplantology 5 Pawinskiego Str, 02-106 Warsaw, POLAND.
Received 2012-4-30; Accepted 2012-6-5; Published 2012-7-1
Abstract
Background: Research over the last twenty years has yielded much insight into pancreatic cancer biology, but it has neither improved diagnostics methods nor the way of treatment. The question remains as to what the critical deciding factor is in making pancreatic cancer such an aggressive disease.
Methods: Pancreatic tumor tissue came from 36 patients. To assess lymphatic vessels color lymphangiography and immunohistochemistry were used. Activity of matrix metalloproteinases was studied with gel and in situ zymography. Expression of growth factors and infiltrating immune cells were investigated using immunohistochemistry.
Results: Our study revealed that the structures that correspond to lymphatic vessels were not observed in tumor center but only at the edge of the tumor. All studied growth factors were present in tumor tissue. We found that the difference in expression between G2 and G3 stage was statistically relevant in cases of c-Met receptor. Inflammatory cells were present around neoplastic glands and also strongly around nerves infiltrated by cancer cells. The number of infiltrating macrophages in tumor tissue was significantly higher in group with metastases to lymph nodes.
Conclusion: We showed two factors that influence pancreatic cancer progression and invasion: c-Met receptors and macrophages infiltrating tumor tissue. Based on our analysis, this indicates that epithelial-mesenchymal transition might be crucial in the progression of pancreatic cancer.
Keywords: pancreatic cancer, growth factors, matrix metalloproteinases, lymphangiogenesis, microenvironment.
Introduction
Pancreatic adenocarcinoma remains one of the deadliest cancers. 1- and 5-year relative survival rates are 25% and 6%, respectively[1]. The surgical intervention remains main possibility of treatment because chemotherapy and radiotherapy has minimal impact on long-term survival. Research over the last twenty years has yielded much insight into pancreatic cancer biology, but it has neither improved diagnostics methods nor the way of treatment. The question remains as to what the critical deciding factor is in making pancreatic cancer such an aggressive disease.
It was shown in recent years that the tumor microenvironment plays a critical role in tumor progression[2]. In pancreatic tumors this microenvironment is particularly heterogeneous. It consists of dense fibrotic stroma with cancer cells, stellate cells, infiltrating inflammatory cells and the remains of the proper structure of the pancreas. These cells are the source of various growth factors: FGF, EGF, PDGF-BB, HGFα as well as proangiogenic factors: VEGF, VEGF C and their receptors. The fibroblasts present in the tumor's tissue are responsible for the production of collagen and fibronectin which increase the chemoresistance of the tumor. Because of abundant fibrotic tissue, the tumor environment is strongly hypoxic[3],[4]. These all factors contribute to the disease's aggressive nature and occurrence of early metastases.
Infiltrating inflammatory cells are a rich source of factors influencing tumor growth, invasion and metastases. Macrophages are of particular interest. Their role in the tumor's environment has been studied in recent years[5]. It is known that they are a rich source of growth factors that stimulate cell proliferation like EGF, PDGF-BB, HGFα and TGF β. Furthermore they also produce matrix metalloproteinase-9 (MMP9) which takes part in various essential processes like tissue remodeling, angiogenesis, epithelial-mesenchymal transition and activating growth factors[6].
As early lymph node metastases are hallmarks of pancreatic cancer lymphatic vessels, they are considered as the main route for the spread of pancreatic cancer. Examining lymphatic vessels makes possible evaluation of lymphangiogenesis inside tumor. In some type of cancer[7],[8] there is a clear evidence linking lymphatic vessels and lymph node metastases, however pancreatic cancer reports on this topic are contradictory[9],[10].
The aim of our study was to investigate three points that may have an influence on the development of the tumor: expression of growth factors, inflammatory cells infiltration and lymphangiogenesis.
Materials and method
Patients and specimen collection
Pancreatic tissue samples were collected from 36 patients who underwent surgical resection due to pancreatic cancer. Tissues were collected based on protocol approved by the Bioethics Committee of Warsaw Medical University. Tumors were classified according to TNM staging and tumor grade (Table 1).
Staging of tumors obtained from patients.
| Parameter | Number of tumors (total n = 36) |
|---|---|
| T stage | |
| T1 | 3 |
| T2 | 6 |
| T3 | 25 |
| T4 | 2 |
| N stage | |
| N0 | 13 |
| N1 | 20 |
| N1a | 2 |
| N1b | 1 |
| M stage | |
| M0 | 33 |
| M1 | 3 |
| Tumor grading | |
| G1 | 4 |
| G2 | 16 |
| G3 | 16 |
For protein isolation, the samples were frozen and stored at -20 °C until they were used. Samples for immunohistochemical analysis with dimensions of 5 x 5 x 5 mm were frozen for 45 seconds in acetone using dry ice at a temperature of -70°C and stored at -80° C.
Color lymphangiography
The analysis was conducted as described previously[11]. In brief, a tissue fragment was injected with 1ml of Paris blue paint suspension in chloroform under low pressure. The injected tissue was fixed in 5% formalin for 1 day and dehydrated for 6 days in turn to 60% 70%, 80%, 96% (two days), 100% ethanol. Then tissue was then placed in methyl salicylate. After at least 3 days it was cut into thick slices and examined under stereomicroscope at 100x magnification.
Immunohistochemistry
Frozen tissue from pancreatic cancer was cryocut into 5µm sections and used for analysis. The Dako REAL™ EnVision™ Detection System, Peroxidase/DAB+, Rabbit/Mouse was used for immunostaining. After being dried in room temperature, the slides were fixed with acetone for 10 min. Then they were incubated for 5 min Dual Endogenous Enzyme Block (Dako, Glostrup, Denmark). The sections were incubated with a proper antibody for a 25 minutes. Afterwards incubation with Dako REAL™ EnVision™/HRP, Rabbit/Mouse (ENV) for 25min at room temperature was followed by a color reaction using Dako REAL™ DAB+ Chromogen for 3 minutes. The slides were counterstained with Mayer's hematoxylin. (Antibodies used for immunohistochemistry are listed in Supplementary Material: Table S1).
Semi-quantitative analysis of immunohistochemical staining
For quantitative evaluation 5 areas were chosen after scanning the tumors sections at low power 40x. These fields were analyzed at 200x magnification using MicroImage software (Olympus, Japan), counting total stained area.
Statistical analysis
A comparison was made with the Mann-Whitney U test. Minimal level of significance was defined as p < 0.05.
Results
Lymphatic vessels in tumor tissue
Color lymphangiography
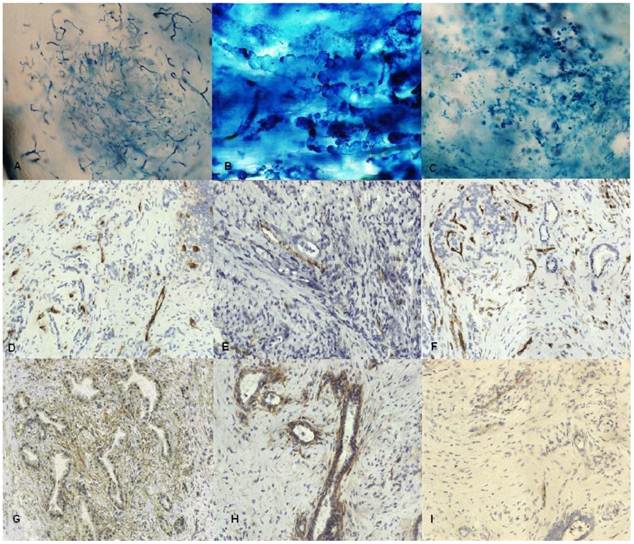
Color lymphangiography pictures of tumor tissue were evaluated. Only occurrence of veins and tiny fluid-filled intercellular microspaces was observed in tissue specimens. The structures that could correspond to lymphatic vessels were not observed in tumor center (Fig 1). Broadened lymphatic vessels tumor could only be observed at the edge of the tumor. Only a few specimens showed blue dye between collagen fibers.
A,B,C color lymphangiography A. Vein vessels in tumor B. Lymphatic vessels from tumor edge C. Microspaces in tumor tissue filled with fluid. Original magnification: x100 D-I Immunostaining for D. FVIII E. Lyve-1 F. CD31 G. podoplanin H. Flt4 I. VEGF C; Original magnification x200.
Immunohistochemistry
To identify lymphatic endothelium we used monoclonal antihuman antibodies against: Prox 1, VEGF C, VEGFR3 (flt4), Lyve1, podoplanin and also chemokines and their receptors CCL21 and CCR7, CXCL13 and CXCR5. To distinguish blood and lymphatic vessels, staining for adhesion molecule CD 31 and v.Willebrandt factor was made. Our study showed that Lyve1 as well as VEGF C, were occasionally present in tumor tissue in structures without lumen (Fig 1). Staining did not show the presence of Prox1, podoplanin was expressed mainly in tumor stroma. VEGFR3 was expressed in the vicinity of cancer nests. Chemokine CCL21 and its receptor CCR7 was produced by cancer cells and some stromal cells, but not by lymphatic endothelium. We did not notice any expression of CXCL13 and its receptor CXCR5.
Growth factors in pancreatic cancer
25 tumor tissues were studied immunohistochemically for expression of growth factors in tumor tissue (Fig. 2). Expression of growth factors was found in all cases. The immunoreactivity of EGF was weak to moderate in cytoplasm of cancer cells. As for EGFR we found its expression to be moderate to strong in cytoplasm of cancer cells and weak in small ductal cells. Strong FGF2 and FGF7 protein expression was observed in the cytoplasm and cytoplasmic membrane of neoplastic cells. Cytoplasmic staining for IGF1 and IGF-IRβ was moderate to strong in cancer cells and positive reaction for IGF-IRβ was also found in stromal cells. PDGF-BB immunoreactivity was moderate to strong in cytoplasm of cancer cells and also in 6 cases we found nuclear staining in cancer cells as well as in infiltrating immune cells. Membranous and cytoplasmic staining for HGFα was strong in tumor cells whereas staining of c-Met was moderate to strong.
Staining for growth factors and their receptors: A. EGF, B. EGFR, C. FGF2, D.FGF7, E.IGF, F. IGFR, G HGFα, H. c-Met, I. PDGF B; original magnification x200
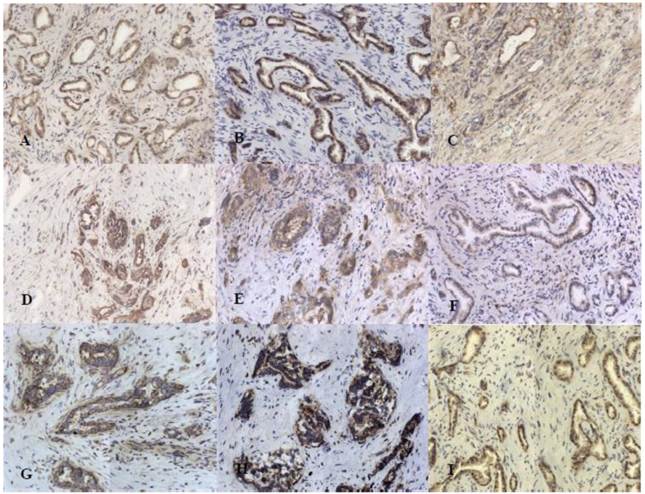
Expression of growth factors was compared against tumor grading and N stage. Using a semiquantitative estimation of all considered growth factors expression, the result of statistical analysis showed that only difference in expression between G2 and G3 group was statistically relevant in case of c-Met receptor (p = 0.033). Estimated mean of expression levels are presented in table 2. Expression of FGF2, FGF7, IGF1 and HGFα was stronger at the G2 stage of tumor. Expression of EGF, EGFR, IGF1-Rβ, PDGF and c-Met was at a higher level in G3 group. In relation to N stage the expression of FGF7, EGFR and IGF1 was lower in N1 group but no difference in expression was statistically significant.
Comparison of expression of growth factors. (NS- not significant, S-significant).
| Antigen | G2 | G3 | Significance | N0 | N1 | Significance |
|---|---|---|---|---|---|---|
| FGF2 | 879,5 | 574,5 | NS | 673,76 | 719 | NS |
| FGF7 | 470 | 371,4 | NS | 414,55 | 389,88 | NS |
| EGF | 270 | 456,17 | NS | 325,04 | 326 | NS |
| EGFR | 344,04 | 533,28 | NS | 427,34 | 362 | NS |
| IGF1 | 509,66 | 435,55 | NS | 442,31 | 406,6 | NS |
| IGFR | 561,55 | 719,98 | NS | 452,15 | 821,82 | NS |
| PDGF | 362,83 | 370 | NS | 348,52 | 398,93 | NS |
| HGFα | 1339,5 | 1014,17 | NS | 1113 | 1225 | NS |
| cMet | 697,56 | 1200,39 | S | 823 | 921,52 | NS |
Infiltrating inflammatory cells
To evaluate cell infiltrates we used monoclonal antibodies against: CD68, HLA II, neutrophil elastase, CD3, CD56. We found numerous lymphocyte and macrophages infiltration. There was also a strong expression of neutrophil elastase. No NK cells infiltration was observed. Inflammatory cells were present around neoplastic glands and also strongly around nerves infiltrated by cancer cells (Fig.3).
Characterization of the inflammatory infiltrate: A. CD3, B.CD68 macrophages, C.elastase, D. CD56; original magnification x200.
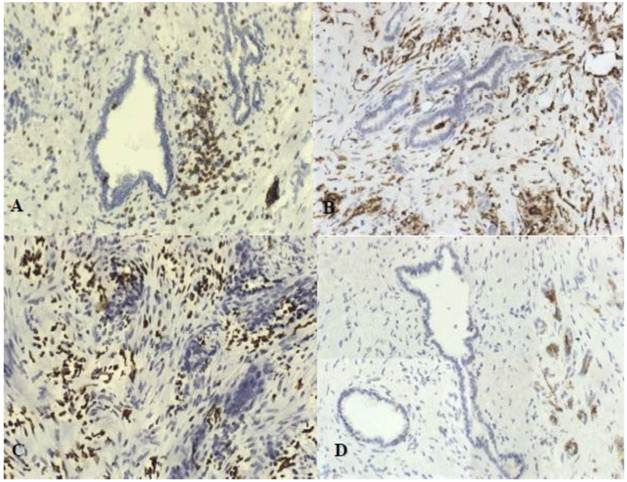
We compared the results due to N stage and we found that the number of macrophages in tumor tissue was significantly higher in group with metastases to lymph nodes (401) than in N0 group (167) (p = 0,0085).
Discussion
Early lymph node metastases are distinguishing mark in pancreatic cancer. The significance of lymphatic vessels and lymphangiogenesis has been widely discussed in recent years[12]. Our research shows that lymphangiogenesis in pancreatic cancer is weak and lymphatic vessels are hardly present in tumor tissue. Also, as was reported previously, expression of VEGF C was weak. Due to strong desmoplastic reaction lymphatic vessels inside tumor are collapsed and incapable of participation in metastases. We found lymphatic vessels only at peripheral tissue surrounding cancer, not in the center of the tumor lesions. In conclusion we claim that dense desmoidal tissue and lack of intratumoral lymphatics preclude lymphatic metastases from the tumor core.
In pancreatic tumors we observed intensive immune cells infiltration. We found that number of macrophages is higher in group with lymph node metastases. This observation supports the statement about participation of macrophages in creating metastases. In pancreatic cancer it was reported previously that macrophages are involved in angiogenesis[13], supporting tumor growth and invasion of cancer cells. They are source of angiogenic factors like VEGF and also MMP9 which degrade extracellular matrix. Important fact is that macrophages can suppress T cell response. Thus, macrophages infiltrating pancreatic tumor are important factor in creating metastases.
Abundant expression of growth factors is typical for pancreatic cancer tissue. As they participate in signaling pathways mediated by receptor tyrosine kinases they might regulate cell proliferation, migration and survival. Previous studies on expression of growth factors provided interesting observation. It was reported that high level of EGF and EGFR correlated with lymph node involvement and distant metastasis as well as reduced median survival[14]. Strong expression FGF2 was told to correlate with advanced tumor stage and increased invasiveness[15]. FGF7 was connected with increased proliferation in pancreatic cancer cells but also in chronic pancreatitis-like lesions adjacent to cancer cells[16]. Also overexpression of IGF1 and IGF1-R was stated in pancreatic cancer and IGF1-R was suggested to be good target for cancer therapy[17],[18]. Only in case of PDGF-BB high levels in blood serum were favorable prognosis[19] and high expression in tumor was related to decreased pancreatic cancer growth[20]. In our research we observed strong expression of growth factors, however we only for c-Met receptor we were able to claim statistically significant difference between G2 and G3 group. No significant difference was observed in relation to lymph node metastases.
Our results indicate that c-Met might be pivotal element in pancreatic cancer. This receptor tyrosine kinase is activated by HGF and physiologically they participate in embryonic development and also during adult life in liver regeneration and wound healing[21]. However their overexpression is observed in pancreatic cancer, even in early stage of carcinogenesis[22].
c-Met activation leads to increased proliferation, enhanced motility and invasion. Therefore we presume that an increase in c-Met expression between G2 and G3 stages indicates more aggressive phenotype which might be related to epithelial-mesenchymal transition. EMT is often considered a first step to metastases as it is related to reduced E-cadherin expression and appearance on N-cadherin[23]. “Cadherin switch” was also observed in pancreatic cancer[24]. Decreased E-cadherin expression was associated with advanced stage and lymph node metastases[25]. Another factor was linked to lymph nodes involvement - presence of macrophages - this was also confirmed by our studies. Our results indicate that the amount of macrophages is significantly higher in group with lymph node metastases, as it was reported previously[26]. Recently it has been shown that macrophages also participate in EMT process[27]. They not only express EMT- inducing cytokines but also MMP9 that cleaves E-cadherin/β-catenin complex. Tan et al. suggested that macrophage MMP9 supports EMT also by disrupting of basement membrane. This is a crucial mechanism for increased motility and invading other tissues.
Apart from HGFα, other growth factors like IGF1, EGF, FGF are also able to induce EMT23. They are all strongly expressed in pancreatic tumor tissue and their overexpression is usually associated with tumor progression and poorer survival[28],[29].
In conclusion, we showed two factors that influence pancreatic cancer progression and invasion: c-Met receptors and macrophages infiltrating tumor tissue. Based on our analysis, this indicates that epithelial-mesenchymal transition might be crucial in the progression of pancreatic cancer. We intend to examine this through further research.
Supplementary Material
Table S1: Antibodies used for immunohistochemistry.
Acknowledgements
We would like to thank prof. Anna Nasierowska-Guttmejer and dr Dorota Łącka for histopathological evaluation of tumors. This study was carried out with support from the ministry of Science and Higher Education nr NN404/0693/33.
Competing Interests
The authors have declared that no competing interest exists.
References
1. American Cancer Society. Cancer Facts & Figures 2010. Atlanta: American Cancer Society. 2010
2. Mueller M.M, Fusenig N.E. Friends or foes - bipolar effects of the tumor stroma in cancer. Nature Rev Cancer. 2004;4:839-849
3. Neoptolemos J.P, Urrutia R.A, Abbruzzese J, Büchler M.W. Pancreatic cancer. Springer. 2010
4. Welsch T, Kleeff J, Friess H. Molecular pathogenesis of pancreatic cancer: advances and challenges. Current Molecular Medicine. 2007;7:504-521
5. Qian BZ, Pollard J.W. Macrophage diversity enhances tumor progression and metastasis. Cell. 2010;141:39-51
6. Pollard J.W. Tumour-educated macrophages promote tumor progression and metastasis. Nat Rev Cancer. 2004;4:71-78
7. Lee K, Park do J, Choe G, Kim HH, Kim WH, Lee HS. Increased intratumoral lymphatic vessel density correlates with lymph node metastasis in early gastric carcinoma. Ann Surg Oncol. 2010;17:73- 80
8. Straume O, Jackson DG, Akslen LA. Independent prognostic impact of lymphatic vessel density and presence of low-grade lymphangiogenesis in cutaneous melanoma. Clin Cancer Res. 2003;9:250- 256
9. Kurahara H, Takao S, Shinchi H, Maemura K, Mataki Y, Sakoda M. et al. Significance of lymphangiogenesis in primary tumor and draining lymph nodes during lymphatic metastasis of pancreatic head cancer. J Surg Oncol. 2010;102:809- 815
10. Sipos B, Kojima M, Tiemann K, Klapper W, Kruse ML, Kalthoff H. et al. Lymphatic spread of ductal pancreatic adenocarcinoma is independent of lymphangiogenesis. J Pathol. 2005;207:301- 312
11. Stańczyk M, Olszewski W.L. et al. Lack of functioning lymphatics and accumulation of tissue fluid/lymph in interstitial "lakes" in colon cancer tissue. Lymphology. 2010;43:158- 167
12. Tammela T, Alitalo K. Lymphangiogenesis: Molecular mechanisms and future promise. Cell. 2010;140:460- 476
13. Esposito I, Menicagli M, Funel N. et al. Inflammatory cells contribute to the generation of an angiogenic phenotype in pancreatic ductal adenocarcinoma. J Clin Pathol. 2004;57:630-6
14. Pryczynicz A, Guzińska-Ustymowicz K, Kemona A, Czyzewska J. Expression of EGF and EGFR strongly correlates with metastasis of pancreatic ductal carcinoma. Anticancer Res. 2008;28:1399-404
15. Yamanaka Y, Friess H, Buchler M. et al. Overexpression of Acidic and Basic Fibroblast Growth Factors in Human Pancreatic Cancer Correlates with Advanced Tumor Stage. Cancer Res. 1993;53:5289-5296
16. Ishiwata T, Friess H, Büchler MW. Characterization of keratinocyte growth factor and receptor expression in human pancreatic cancer. Am J Pathol. 1998;153:213-22
17. Bergmann U, Funatomi H, Yokoyama M. et al. Insulin-like Growth Factor I Overexpression in Human Pancreatic Cancer: Evidence for Autocrine and Paracrine Roles. Cancer Res. 1995;55:2007-2011
18. Golan T, Javle M. Targeting the insulin growth factor pathway in gastrointestinal cancers. Oncology (Williston Park). 2011;25:518-26
19. Rahbari NN, Schmidt T, Falk CS. et al. Expression and prognostic value of circulating angiogenic cytokines in pancreatic cancer. BMC Cancer. 2011;11:286
20. McCarty MF, Somcio RJ, Stoeltzing O. Overexpression of PDGF-BB decreases colorectal and pancreatic cancer growth by increasing tumor pericyte content. J Clin Invest. 2007;117:2114-22
21. Gentile A, Trusolino L, Comoglio PM. The Met tyrosine kinase receptor in development and cancer. Can Met Rev. 2008;27:85- 94
22. Yu J, Ohuchida K, Mizumoto K, Ishikawa N, Ogura Y, Yamada D. et al. Overexpression of c-met in the early stage of pancreatic carcinogenesis; altered expression is not sufficient for progression from chronic pancreatitis to pancreatic cancer. World J Gastroenterol. 2006;12:3878- 3882
23. Thiery J. P: Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002;2:422- 454
24. Nakajima S, Doi R, Toyoda E, Tsuji S, Wada M, Koizumi M, Tulachan SS. et al. N-cadherin expression and epithelial-mesenchymal transition in pancreatic carcinoma. Clin Cancer Res. 2004;10:4125- 4133
25. Joo YE, Rew J, Park C, Kim S. Expression of E-cadherin, alpha- and beta-catenins in patients with pancreatic adenocarcinoma. Pancreatology. 2002;2:129- 137
26. Kurahara H. Significance of M2-polarized tumor-associated macrophage in pancreatic cancer. J Surg Res. 2011;167:e211- 9
27. Tan TK, Zheng G, Hsu TT, Wang Y, Lee VW, Tian X, Wang Y. et al. Macrophage matrix metalloproteinase-9 mediates epithelial-mesenchymal transition in vitro in murine renal tubular cells. Am J Pathol. 2010;176:1256- 70
28. Ozawa F, Friess H, Tempia-Caliera A, Kleeff J, Büchler M W. Growth factors and their receptors in pancreatic cancer. Teratog Carcinog Mutagen. 2001;21:27-44
29. Mihaljevic A.L, Michalski Ch, Friess H, Kleeff J. Molecular mechanism of pancreatic cancer—understanding proliferation, invasion, and metastasis. Langenbecks Arch Surg. 2010;395:295-308
Author contact
![]() Corresponding author: Phone (+4822) 6086521, fax (+4822) 6685334, email: kgardiancom.
Corresponding author: Phone (+4822) 6086521, fax (+4822) 6685334, email: kgardiancom.