Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2019; 10(8):1800-1807. doi:10.7150/jca.29889 This issue Cite
Research Paper
CUEDC2 Contributes to Cisplatin-Based Chemotherapy Resistance in Ovarian Serious Carcinoma by Regulating p38 MAPK Signaling
Aichun Wang1,2*, Jinhang Li1*, Tao Zhou3, Tao Li3, Hong Cai3, Huaiyin Shi1 ![]() , Aijun Liu1
, Aijun Liu1 ![]()
1. Department of Pathology, People's Liberation Army General Hospital, Beijing, 100853, China
2. Department of Pathology, Haidian Maternal & Children Health Hospital, Beijing, 100080, China
3. National Center of Biomedical Analysis, Institute of Basic Medical Sciences, Beijing, 100850, China
*These authors contributed equally to this project.
Received 2018-9-12; Accepted 2019-2-7; Published 2019-4-21
Abstract
Chemoresistance remains an obstacle to the successful treatment of ovarian carcinoma. CUE domain-containing 2 (CUEDC2) plays critical roles in tumor genesis and overexpresses in many solid cancers, including ovarian serous carcinoma. In previous study, we found that overexpression of CUEDC2 might be a promising biomarker to evaluate the progression and to predict likely relapse of serous ovarian carcinoma. In present study, we found that higher expression of CUEDC2 was associated with higher resistance to cisplatin. The overall survival (OS) and disease-free survival time (DFS) of patients with cisplatin resistant was shorter than that of those with cisplatin sensitive, respectively, and the cisplatin sensitivity was independent predictor of a shorter OS time and DFS time. Knockdown of CUEDC2 by small interfering RNA enhanced the cisplatin sensitivity of serous ovarian carcinoma cells in SKOV3 cell lines. Furthermore, the phosphorylation of p38 MAPK were obviously increased after CUEDC2 knockdown, while p38 MAPK signaling contributes to cell growth and cell apoptosis. Our data suggest that CUEDC2 takes part in cisplatin-based chemotherapy resistance by regulating p38 MAPK signaling. And CUEDC2 is a promising biomarker and therapeutic target of cisplatin resistance in ovarian serous carcinoma.
Keywords: CUEDC2, ovarian carcinoma, chemoresistance, cisplatin, p38 MAPK
Introduction
Ovarian carcinoma is the most lethal gynecologic malignancy leading a cause of mortality with 239,000 new cases and 152,000 deaths worldwide in 2012, accounting for 3.6% of all cancer deaths in women [1]. There were 521, 000 new cases of ovarian carcinoma diagnosed in the mainland of China, and 5-year survival rate of patients diagnosed with advanced stage was less than 20%, even with reduction and chemotherapy [2]. Chemotherapy combined with tumor reduction has been confirmed as the first-line therapeutic protocol. Cisplatin is the most widely used crosslinking drug for killing ovarian cancer cells, and cisplatin-based chemotherapy improves patient survival [3]. Unfortunately, although the majority of ovarian carcinoma was initially sensitive to chemotherapy, long-term administration of cisplatin developed chemotherapeutic resistance. Chemoresistance remains a bottleneck problem in the course of ovarian carcinoma treatment [4]. The elucidation of drug resistant mechanisms is insufficient to overcome clinical resistance. Therefore, to explore drug resistant mechanisms and enhance the chemotherapeutic drug sensitivity is an urgent problem to be solved [5].
CUE domain-containing 2 (CUEDC2), a CUE domain-containing protein, is a small and moderately conserved ubiquitin-binding domain of about 40 amino acids. CUEDC2 has been found in a variety of eukaryotic proteins and plays critical roles in many biological processes, such as cell cycle, inflammation and tumor genesis [6-7]. Gao et al demonstrated that high levels of CUEDC2 might result in earlier inactivation of the anaphase-promoting complex or cyclosome (APC/C), eventually resulting in chromosome missegregation and aneuploidy, and then promote tumor formation [8]. What is more, CUEDC2 overexpressed in many solid cancers such as breast cancer, ovarian cancer, and brain cancer. Pan et al demonstrated that CUEDC2 protein could confer endocrine resistance in breast cancer through the ubiquitin-proteasome pathway [9]. Recent study demonstrated that sinomenine inhibited breast cancer cell invasion and migration by suppressing NF-κB activation mediated by IL-4/miR-324-5p/CUEDC2 axis [10]. Zhang et al suggested that the expression level of CUEDC2 had an inverse correlation with imatinib resistance and activity of NF-κB signaling pathway in chronic myeloid leukemic cells [11].
The role of CUEDC2 in ovarian serous carcinoma remains unclear. In previous study, we found CUEDC2 overexpression was correlated with advanced FIGO stage and recurrence, which implied that CUEDC2 might be a promising biomarker to evaluate the progression and predict likely relapse of serous ovarian carcinoma [12]. Moreover, clinical studies suggested that incidence of cisplatin resistance was increasing with tumor stage advancing, which implicated that CUEDC2 might take part in cisplatin resistance during ovarian serous carcinoma treatment.
MAPKs are essential components of the signaling transduction mechanism, which being highly involved in cell growth, differentiation and cell apoptosis [13]. Studies have suggested apoptotic stimuli to be transmitted to caspases through the activation of MAPKs, such as p38 MAPK and JNK [14-15]. The p38 MAPK pathway is involved in cancer cell apoptosis and induced by several chemotherapeutic drugs [16]. The loss of the capacity to activate p38 MAPK in response to cisplatin treatment was also reported to be a potential mechanism of chemoresistance [17].
In this study, we detected the expression of CUEDC2 in ovarian serous carcinoma tissues and analyzed its correlation with sensitivity to chemotherapy agents among ovarian serous carcinoma patients. We found that cisplatin resistance cases all presented CUEDC2 positive expression, while 73.2% of sensitivity cases. Furthermore, we analyzed the effect of CUEDC2 inhibition on chemosensitivity and cell proliferation by down-regulating CUEDC2 expression with siRNA in ovarian cancer cells, in order to determine whether CUEDC2 is key factor in cisplatin-based chemotherapy of ovarian serous carcinoma.
Materials and Methods
Reagents
Human ovarian serous carcinoma cell line SKOV3 was purchased from the ATCC cell bank. Anti-human CUEDC2 antibody was kindly donated by Professor Xuemin Zhang (Chinese National Center of Biomedical Analysis, Institute of Basic Medical Sciences). Anti-human Tubulin and anti-human p-p38 MAPK were purchased from Cell Signaling Technology (Boston, USA). Fetal bovine serum was purchased from Gib Co. (New York, USA), CellTiter 96® AQueous One Solution Cell Proliferation Assay from Promega Co. (Madison, USA), and Lipofectamine RNAiMax Reagents from Invitrogen Co. (Carlsbad, USA), respectively. Cisplatin was provided by Qilu Pharmaceutical Co., Ltd (Ji'nan, China).
Samples selection
We previously reported the overexpression of CUEDC2 was observed in 59.4% (60/101) of ovarian serous carcinoma tissue samples and correlated with histopathological grade, patient age at diagnosis, FIGO stage and recurrence. Further analysis showed that the major cases of cisplatin resistance presented CUEDC2 overexpression. To conform whether CUEDC2 affects cisplatin chemosensitivity, we collected 62 ovarian serous carcinoma cases who first received cisplatin-based chemotherapy from January 2007 to April 2012 at the Chinese People's Liberation Army General Hospital. Cisplatin sensitivity was defined by the response to cisplatin-based chemotherapy or a progression-free interval >6 months off treatment. Patients were considered to have progressed during treatment if they had a stable disease in response to initial cisplatin-based therapy, and those who relapsed within 6 months were considered to have cisplatin-resistance disease. Patients were excluded if 1) undergoing neoadjuvant chemotherapy before the surgical operation, 2) tumor tissue paraffin blocks were unavailable or inadequate, or 3) with other synchronous or metastatic tumors to the ovary. Tissue specimens were formalin fixed and paraffin embedded (FFPE).
Ethical statement
The study using clinical samples was approved by Institutional Review Board at the Chinese PLA General Hospital Ethics Board. Informed consents were obtained from all subjects or their relatives.
Immunohistochemistry (IHC) staining
We did immunohistochemistry staining on all tumor samples with anti-CUEDC2 antibody. Sections from FFPE tissues were used for immunohistochemical staining according to a standard method and German semi-quantitative scoring system was used as we reported previously [11]. Each specimen was assigned a score according to the intensity of cytoplasmic staining (no staining = 0; weak staining = 1, moderate staining = 2, strong staining = 3) and the extent of stained cells (0% = 0, 1-24% = 1, 25-49% = 2, 50-74% = 3, 75-100% = 4). The final immunoreactive score was determined by multiplying the intensity score with the extent of score of stained cells, ranging from 0 (the minimum score) to 12 (the maximum score). We defined 0 score as negative, 1-6 score as weak positive expression and ≥8 score as strong positive expression, respectively.
Cell culture and siRNA transfection
Human ovarian serous carcinoma cell line SKOV3 were cultured in DMEM containing 1% penicillin (100IU/ml) and streptomycin (100μg/ml), supplemented with and 10% fetal bovine serum (FBS) at 37 °C in a humidified atmosphere of 5% CO2. For cisplatin treatment experiments, cells were cultured in medium containing phenol red-free DMEM supplemented with 5% charcoal-filtered fetal bovine serum.
The SKOV3 cells were seeded into 96-well plates at 6×103 cells/well and transfected with siCUEDC2 (target sequence: 5'- CUCAGCGCCAGUUGCCUCAUCUUGG -3') or scrambled siRNA using LipofectamineTM2000 (Invitrogen) according to the manufacturer's instruction. The cells were harvested after 72h later. Scrambled siRNA oligos (si-control) was used as a negative control.
Measurement of cell viability
The viability of SKOV3 cells were examined by MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetra-zolium bromide] assay according to manufacturer's instruction. Briefly, cells were seeded on 96-well microtitre plates at a density of 6× 103 cells per well. Cells were cultured for 24h, 48h, 72h and 96h, respectively. After indicated time of incubation, CellTiter 96® AQueous One Solution Reagent was added. Then the cells were incubated at 37◦C for an additional 4h, and triple lysate (sodium dodecyl sulfate/isobutanol/HCl) was added to each well. The absorbance of formazan product was measured by Versa Max tunable microplate reader (Molecular Devices, Sunnyvale, CA, USA) at 490nm. Each experiment was repeated at least thrice in quadruplicate.
Cisplatin sensitivity assays
After 48h transfection with siRNA molecules on SKOV3 cells, cisplatin cytotoxicity was measured using the MTT assay. Cells were plated at 6×103 cells/well in 96-well plates in triplicate for each dose. Cisplatin was added with the desired drug treatment concentrations ranging from 0 to 64μM and incubated for 72h. Formazan products were solubilized with DMSO, and the optical densities were measured at 490nm. IC50 was used as the measure of relative cytotoxicity.
Western blot
Total protein was extracted using RIPA lysis buffer with 1mM PMSF (Sigma, USA) and complete protease inhibitor cocktail (Roche). Protein concentration was determined by the BCA assay (Beijing, China) according to the manufacturer's instructions. The whole-cell lysates mixed with loading buffer were heat-denatured at 100 ◦C for 10min before being run on 8-12% gradient SDS-PAGE. After SDS-PAGE, the proteins were electrotransferred onto PVDF membranes and were blotted with primary antibody and secondary antibody. Then the samples were detected with enhanced chemiluminescence.
Statistical analysis
All data were processed using SPSS 19.0 statistical software. The correlation between CUEDC2 expression and cisplatin resistance was analyzed by chi-square test. The difference between two subgroups was analyzed by two independent samples t-test. Survival curves were constructed with the Kaplan-Meier product-limit method and compared by the log-rank test. To evaluate the independent prognostic factors associated with survival, multivariate Cox proportional hazards regression analysis was used. All statistical tests were two-sided and P values < 0.05 were considered to be statistically significant.
Results
The positive expression of CUEDC2 increased with the histopathological grading and the rates of cisplatin resistance correlated with the CUEDC2 expression in serous ovarian carcinoma patients
The mean age of 62 patients with cisplatin-based chemotherapy at the time of diagnosis was 51.7 years old (range 21-74 y). Forty seven cases were categorized into HGSC and 15 cases were LGSC. Fourteen out of 62 cases were in early stage(FIGO Ⅰ-Ⅱ) and 48 cases in advanced stage (FIGO Ⅲ-Ⅳ).Among 62 cases, there were 21 cases presenting cisplatin resistance and relapsed within 6 months. A total of 57 cases had followed up records were included in data analysis. The mean disease-free survival time (DFS) of 57 cases was 24 months and mean overall survival (OS) time was 70 months.
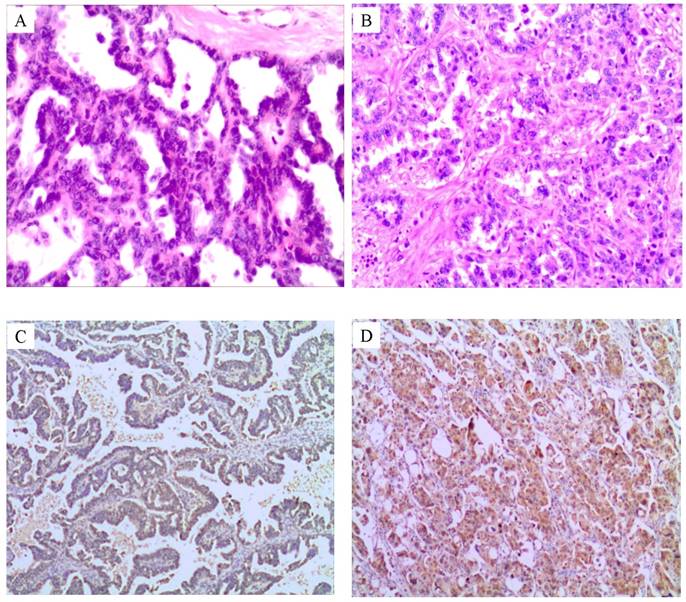
As shown in Fig.1, the positive expression of CUEDC2 increased with the histopathological grading (Fig.1 A-D). Twenty one cases with cisplatin resistance all presented CUEDC2 positive expression (100%, 21/21), 30 out of 41 cases presented positive expression in cisplatin sensitivity subgroup (73.1%, 30/41), which had statistical significance (P =0.011) as shown in Table 1.
The positive expression of CUEDC2 increased with the histopathological grading in serous ovarian carcinoma. A. Low-grade ovarian serous carcinoma, papillary structures with uniform homogeneous epithelial cells, mild to moderate nuclear atypia, and rare necrosis. HE×100. B. High-grade serous carcinoma,with marked nuclear atypia and common mitosis. HE×100. C. CUEDC2 weakly immunostained with cytoplasm in light yellow color in LGSC subgroup. D. CUEDC2 strongly immunostained with cytoplasm in yellow or brown color in HGSC subgroup.
Correlation of clinicopathological variables with CUEDC2 expression in patients with cisplatin-based chemotherapy of serous ovarian carcinoma
Variable | CUEDC2 | P value (Fisher exact test) | ||
|---|---|---|---|---|
| negative | positive | n | ||
| Age | ||||
| ≤50 | 6 | 20 | 26 | 0.502 |
| >50 | 5 | 31 | 36 | |
| Grading | ||||
| LGSC | 6 | 9 | 15 | 0.018 |
| HGSC | 5 | 42 | 47 | |
| FIGO stage | ||||
| Ⅰ-Ⅱ | 4 | 11 | 15 | 0.437 |
| Ⅲ-Ⅳ | 7 | 40 | 47 | |
| Reaction to cisplatin | ||||
| Sensitivity | 11 | 30 | 41 | 0.011 |
| Resistance | 0 | 21 | 21 | |
The OS time and DFS time of patients with cisplatin resistant was shorter than that of those with cisplatin sensitive, respectively, and the cisplatin sensitivity was independent predictor of shorter OS time and DFS time for patients
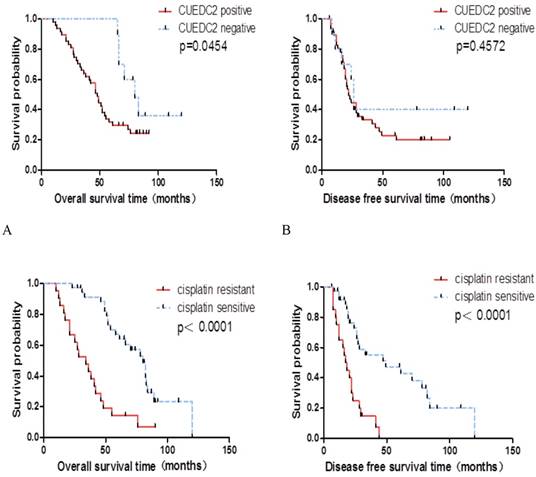
In univariate survival analysis, the association of CUEDC2 expression and cisplatin chemosensitivity achieved statistical significance for OS time, and the DFS time of CUEDC2 negative subgroup was slighter longer than that of CUEDC2 positive subgroup, but no statistical significance was attained (Table 2). Furthermore, the OS time and DFS time of the cisplatin resistant subgroup was shorter than that of the sensitive subgroup, respectively (Fig.2).
Univariate survival analysis of prognostic factors and CUEDC2 expression in patients with cisplatin-based chemotherapy of serous ovarian carcinoma
| Variable | OS time (month) | P value | DFS time (month) | P value | |
|---|---|---|---|---|---|
| CUEDC2 | Negative | 82.7±18.74 | <0.001 | 44.50±41.71 | 0.180 |
| Positive | 49.11±24.12 | 31.45±23.80 | |||
| Cisplatin | Sensitivity | 65.92±22.83 | 0.001 | 42.44±30.96 | 0.001 |
| Resistance | 36.29±21.62 | 18.81±10.852 | |||
| Grading | LGSC | 59.33±34.51 | 0.518 | 40.42±39.42 | 0.354 |
| HGSC | 53.84±24.24 | 31.96±24.09 | |||
| FIGO Stage | Ⅰ-Ⅱ | 68.92±27.15 | 0.030 | 44.85±35.071 | 0.102 |
| Ⅲ-Ⅳ | 50.89±25.11 | 30.45±24.811 | |||
| Age | ≤50 | 61.08±30.43 | 0.140 | 39.96±32.28 | 0.151 |
| >50 | 50.58±22.63 | 29.21±23.039 | |||
The OS time and DFS time of the cisplatin resistant subgroup was shorter than that of the sensitive subgroup, respectively. A. OS time of CUEDC2-negtive patients versus CUEDC2 -positive patients, there was statistically significant difference (P=0.0454) B. DFS time of CUEDC2-negtive patients versus CUEDC2 -positive patients, there was no statistically significant difference (P=0.4572). C. OS time of cisplatin-resistant patients versus cisplatin-sensitive patients, there was statistically significant difference (P<0.001). D. DFS time of cisplatin-resistant patients versus cisplatin-sensitive patients, there was statistically significant difference (P<0.001)
Multivariate survival analysis based on the Cox proportional hazards regression model was performed on all parameters. The result of this analysis was shown in Table 3. It can be seen that the cisplatin sensitivity was independent predictor of shorter OS time and DFS time for patients undergoing curative resection (P=0.002 and P=0.003), meanwhile FIGO stage was a potential independent predictor of shorter OS time (P=0.051).
Independent predictors of survival in patients with cisplatin-based chemotherapy of serous ovarian carcinoma
| Variable | OS time | DFS time | ||||
|---|---|---|---|---|---|---|
| HR (95%CI) | P value | HR (95%CI) | P value | |||
| CUEDC2 | Positive | 0.569 (0.207,1.563) | 0.274 | 0.965 (0.353,2.642) | 0.945 | |
| Negative | ||||||
| Cisplatin | Cisplatin | 0.341 (0.172,0.676) | 0.002 | 0.345 (0.170,0.700) | 0.003 | |
| Sensitivity | ||||||
| FIGO | Ⅲ—Ⅳ | 0.385 (0.148,1.004) | 0.051 | 0.502 (0.192,1.307) | 0.158 | |
| Ⅰ—Ⅱ | ||||||
CUEDC2 siRNA effectively reduces CUEDC2 protein levels in ovarian cancer SKOV3 cells
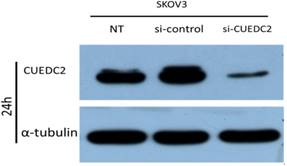
To explore the possible effects of CUEDC2 on cisplatin sensitivity, we examined whether CUEDC2 affects cisplatin sensitivity in ovarian serous carcinoma cell. The CUEDC2 specific siRNA plasmids were used to downregulate CUEDC2 expression in SKOV3 cells. As expected, stable transfection experiments showed siRNA could significantly inhibit CUEDC2 expression in SKOV3 cells in protein level (Fig.3).
CUEDC2 was knocked down in SKOV3 cells by using siRNA (50nM) and analyzed by western blot analysis at 24h.
MTT assays were performed to determine whether the knockdown of cellular CUEDC2 level inhibit cell proliferation. The results showed that CUEDC2 siRNA could suppress cell proliferation more than that of the si-control lines after a certain concentration of cisplatin (8μM) were added to cell lines. The inhibition occurred at 24h and became more significant after that point as showed in Fig.4A.
Knocking down of CUEDC2 enhanced the cisplatin sensitivity of serous ovarian carcinoma cells. A. Cell proliferation curve of si-control compared with si-CUEDC2 subgroup with concentration of cisplatin (8µM) DDP (a: p<0.05). B. Cisplatin inhibited the proliferation of SKOV3 cell in si-CUEDC2 compared with si-control subgroup (a: P<0.05,b: P<0.01)
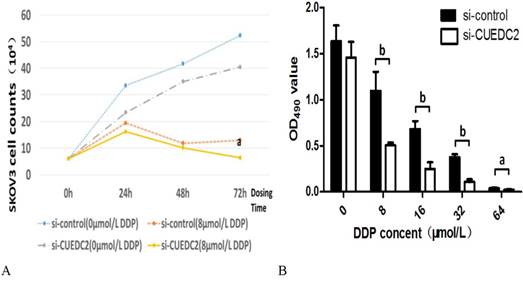
In order to further study the effect of CUEDC2 expression on cisplatin sensitivity, different concentrations of cisplatin were added to the si-control and si-CUEDC2 subgroup cell lines. After 72 hours of incubation, the inhibitory effect on the proliferation of ovarian cancer cells was gradually enhanced with increasing drug concentration (P<0.05); Compared with the si-control subgroup, cisplatin inhibited the cell proliferation more significantly in si-CUEDC2 subgroup as shown in Fig.4B(P<0.05).
Taken together, these results demonstrate that siRNA-mediated CUEDC2 inhibition could result in an obvious decrease in cell proliferation and enhance the cisplatin sensitivity of serous ovarian cancer cells.
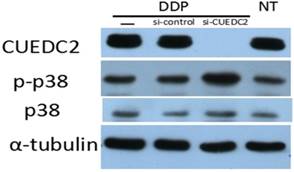
CUEDC2 contributes to cisplatin-based chemotherapy resistance by inhibiting the phosphorylation of p38 MAPK
MAPKs are essential components of the signaling transduction mechanism, while being highly involved in cell growth and cell apoptosis [13]. Studies have suggested apoptotic stimuli to be transmitted to caspases through the activation of p38 MAPK pathway. To rule out the correlation between CUEDC2 expression and p38 signaling pathway activity, we investigated the activity of p38 after CUEDC2 was knocked down in SKOV3 cells. The results showed that phosphorylation of p38 was obviously increased (Fig.5). This finding suggested CUEDC2 high expression could inhibit cisplatin- induced cell death and promote cell proliferation by blocking p38 phosphorylation.
Knocking down of CUEDC2 enhanced p38 MAPK signal activation. Lane —: Cells were treated with 8µM cisplatin (DDP); Lane si-control: Cells of si-control subgroup were treated with 8µM cisplatin (DDP); Lane si-CUEDC2: Cells of si-CUEDC2 subgroup were treated with 8µM cisplatin (DDP); Lane NT: None treat. Tubulin was used as loading control
Discussion
Resistance to chemotherapies is an obstacle to the successful treatment of ovarian carcinoma [19]. Therefore, the identification of new markers with predictive value in response to cisplatin treatment will be helpful for improving the efficacy and minimizing the side effects of cisplatin [20].
In the study, we found that higher expression of CUEDC2 was associated with higher resistance to cisplatin, which may account for the OS time and DFS time of the cisplatin resistant subgroup were shorter than those of the sensitive subgroup. Considering the effect of histopathological grade to cisplatin, we analyzed the relationship of CUEDC2 expression and the cisplatin sensitivity in LGSC and HGSC group, respectively. The results showed that the cases with CUEDC2-positive were more likely to resistant than those with CUEDC2-negative in each group, but there was no statistical significance which may be due to the insufficient samples.
By down-regulating CUEDC2 expression, we found that CUEDC2 deficiency could result in an obvious decrease in cell proliferation and enhance the cisplatin sensitivity of serous ovarian carcinoma cells. Also, the results showed that CUEDC2 contributed to the sensitivity of serous ovarian carcinoma cells to cisplatin. Then we found the phosphorylation of p38MAPK were obviously increased after CUEDC2 was knocked down in SKOV3 cells. Our results implicated that CUEDC2 overexpression could inhibit cisplatin-induced cell death and promote cell proliferation by inhibiting p38 activation.
As a key player in the response to chemotherapy, the p38MAPK pathway is involved in cancer cell apoptosis and could be activated by several chemotherapeutic drugs [16]. The role as a determinant of resistance to cisplatin was proposed mainly based on the inhibition of p38MAPK in several experimental models [21-22]. Hyperactivation of p38 MAPK has been correlated with cisplatin resistance [23]. There have been report showed that protein phosphatase 1 (PP1) regulates the phosphorylation state of p38, and that p38 phosphorylation can be enhanced by PP1 inhibitors [24]. And CUEDC2 has been demonstrated to recruit PP1 and repressed activation of the kinase IKK [25]. On the other side, NF-κB has emerged as a promising anti-cancer target to overcome resistance to chemotherapy and activated NF-kB has also been identified as a key mechanism of cisplatin resistance [26]. Report has demonstrated that inactivating mutations in the MAPK pathway could lead to signaling via the NF-kB pathway in response to cisplatin treatment [27]. Thereby, detailed mechanism of how CUEDC2 regulating P38MAPK signaling activation is unknown and require further investigation.
In summary, CUEDC2 down-regulation can enhance the drug sensitivity to cisplatin in ovarian serous carcinoma through p38MAPK signaling pathway. It appears that CUEDC2 might represent a novel therapy strategy and serve as an indicator for disease progression and therapy response in ovarian serous carcinoma. Further detailed investigation of the association of CUEDC2 and cisplatin resistance, as well as the mechanism of adjusting the CUEDC2 expression in ovarian serous carcinoma should be performed. In conclusion, our study raises the possibility that CUEDC2 acts as a promising biomarker and therapeutic target of cisplatin resistance in ovarian serous carcinoma.
Acknowledgements
We thank Professor Xuemin Zhang for his generous donation of the antibody to CUEDC2.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M. et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136:359-86
2. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F. et al. Cancer statistics in China,2015. CA Cancer J Clin. 2016;66:115-32
3. Galluzzi L, Senovilla L, Vitale I, Michels J, Martins I, Kepp O. et al. Molecular mechanisms of cisplatin resistance. Oncogene. 2012;31:1869
4. Yan XD, Li M, Yuan Y, Mao N, Pan LY. Biological comparison of ovarian cancer resistant cell lines to cisplatin and Taxol by two different administrations. Oncol Rep. 2007;17:1163-69
5. Livney YD, Assaraf YG. Rationally designed nanovehicles to overcome cancer chemoresistance. Adv Drug Deliv Rev. 2013;65:1716-30
6. Man J, Zhang X. CUEDC2: an emerging key player in inflammation and tumorigenesis. Protein Cell. 2011;2:699-703
7. Zhong X, Tian S, Zhang X, Diao X, Dong F, Yang J. et al. CUE domain-containing protein 2 promotes the Warburg effect and tumorigenesis. Embo Reports. 2017:809-25
8. Gao YF, Li T, Chang Y, Wang YB, Zhang WN, Li WH. et al. Cdk1-phosphorylated CUEDC2 promotes spindle checkpoint inactivation and chromosomal instability. Nat Cell Biol. 2011;13:924-33
9. Pan X, Zhou T, Tai YH, Wang C, Zhao J, Cao Y. et al. Elevated expression of CUEDC2 protein confers endocrine resistance in breast cancer. Nat Med. 2011;7:708-14
10. Song L, Liu D, Zhao Y, He J, Kang H, Dai Z. et al. Sinomenine inhibits breast cancer cell invasion and migration by suppressing NF-κB activation mediated by IL-4/miR-324-5p/CUEDC2 axis. Biochem Biophys Res Commun. 2015;464:705-10
11. Zhang H, Chang G, Wang J, Lin Y, Ma L, Pang T. CUEDC2 sensitizes chronic myeloid leukemic cells to imatinib treatment. Leuk Res. 2013;37:1583-91
12. Wang A, Guo C, Sun Y, Lu L, Wang Y, Wang Q. et al. Overexpression of CUEDC2 predicts poor prognosis in ovarian serous carcinomas. J Cancer. 2015;6:542-47
13. Ono K, Han J. The p38 signal transduction pathway: activation and function. Cell Signal. 2000;12:1-13
14. Park SJ, Kim IS. The role of p38 MAPK activation in auranofin-induced apoptosis of human promyelocytic leukaemia HL-60 cells. Br J Pharmacol. 2005;146:506-13
15. Wagner EF, Nebreda AR. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat Rev Cancer. 2009;9:537-49
16. Zhang B, Wang X, Cai F, Chen W, Loesch U, Zhong XY. Antitumor properties of salinomycin on cisplatin-resistant human ovarian cancer cells in vitro and in vivo: Involvement of p38 MAPK activation. Oncology Report. 2013;29:1371-78
17. Brozovic A, Fritz G, Christmann M, Zisowsky J, Jaehde U, Osmak M. et al. Long-term activation of SAPK/JNK, p38 kinase and fas-L expression by cisplatin is attenuated in human carcinoma cells that acquired drug resistance. Int J Cancer. 2004;112:974-85
18. Zhang WN, Wang L, Wang Q, Luo X, Fang DF, Chen Y. et al. CUEDC2 (CUE Domain-containing 2) and SOCS3 (Suppressors of Cytokine Signaling 3) Cooperate to Negatively Regulate Janus Kinase 1/Signal Transducers and Activators of Transcription 3 Signaling. J Biol Chem. 2012;287:382-92
19. Lawrie TA, Winter-Roach BA, Heus P, Kitchener HC. Adjuvant (post-surgery) chemotherapy for early stage epithelial ovarian cancer. Cochrane Database Syst Rev. 2015;17:CD004706
20. Wang Y, Wang H, Song T, Zou Y, Jiang J, Fang L. et al. HOTAIR is a potential target for the treatment of cisplatin-resistant ovarian cancer. Mol Med Rep. 2015;12:2211-16
21. Cuadrado A, Lafarga V, Cheung PC, Dolado I, Llanos S, Cohen P. et al. A new p38 MAP kinase-regulated transcriptional coactivator that stimulates p53-dependent apoptosis. EMBO J. 2007;26:2115-26
22. St St Germain C, Niknejad N, Ma L, Garbuio K, Hai T, Dimitroulakos J. Cisplatin induces cytotoxicity through the mitogen-activated protein kinase pathways and activating transcription factor 3. Neoplasia. 2010;12:527-38
23. Galan-Moya EM, de la Cruz-Morcillo MA, Llanos Valero M, Callejas-Valera JL, Melgar-Rojas P, Hernadez Losa J. et al. Balance between MKK6 and MKK3 mediates p38 MAPK associated resistance to cisplatin in NSCLC. PLoS One. 2011;6:e28406
24. Samuvel DJ, Jayanthi LD, Bhat NR, Ramamoorthy S. A role for p38 mitogen-activated protein kinase in the regulation of the serotonin transporter: evidence for distinct cellular mechanisms involved in transporter surface expression. J Neurosci. 2005;25:29-41
25. Li HY, Liu H, Wang CH, Zhang JY, Man JH, Gao YF. et al. Deactivation of the kinase IKK by CUEDC2 through recruitment of the phosphatase PP1. Nat Immunol. 2008;9:533-41
26. Godwin P, Baird AM, Heavey S, Barr MP, O'Byrne KJ, Gately K. Targeting nuclear factor -kappa B to overcome resistance to chemotherapy. Front Oncol. 2013;3:120
27. Sanchez-Perez I, Benitah SA, Martinez-Gomariz M, Lacal JC, Perona R. Cell stress and MEKK1-mediated c-Jun activation modulate NFkappaB activity and cell viability. Mol Biol Cell. 2002;13:2933-45
Author contact
![]() Corresponding author: Aijun Liu (liuaijuncom.cn) or Huaiyi Shi (shihuaiyincom), Department of Pathology, People's Liberation Army General Hospital, Beijing, 100853, China.
Corresponding author: Aijun Liu (liuaijuncom.cn) or Huaiyi Shi (shihuaiyincom), Department of Pathology, People's Liberation Army General Hospital, Beijing, 100853, China.