Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2026; 17(6):1229-1247. doi:10.7150/jca.134550 This issue Cite
Review
Emerging Oncolytic Viruses: Beyond Adenoviruses and Herpes Simplex Virus
Santosh Chokkakula1#, Chengliang Yin2#, Bing Yang3,4#, Siomui Chong5,6, Balaji Pathakumari7, Yi-quan Zhang4, Pangi Vijaya Nirmala8, Khin Wee Lai2 ![]() , Bommireddy Naveen9
, Bommireddy Naveen9 ![]() , Quan Tang10
, Quan Tang10 ![]()
1. Department of Microbiology, Chungbuk National University College of Medicine and Medical Research Institute, Cheongju, Chungbuk, 28644, South Korea.
2. Department of Biomedical Engineering, Faculty of Engineering, Universiti Malaya, Kuala Lumpur, 50603, Malaysia.
3. Department of Medical and Nursing Science, International College, Krirk University, Bangkok 10220, Thailand.
4. Department of Cell Biology, College of Basic Medical Sciences, Tianjin Medical University, Tianjin, China.
5. Department of Dermatology, The University of Hong Kong-Shenzhen Hospital, Shenzhen, 518053, China.
6. Department of Dermatology, The First Affiliated Hospital of Jinan University & Jinan University Institute of Dermatology, Guangzhou, 510630, China.
7. Division of Pulmonary and Critical Care Medicine, Department of Medicine, Mayo Clinic, Rochester, MN, 55905, USA.
8. School of Life & Health Sciences Adikavi Nannaya University Rajahmundry, Andhra Pradesh India, 533296, India.
9. Department of Chemical and Biological Engineering, Gachon University, Seongnam-Si, Gyeonggi-do, Republic of Korea.
10. Research Laboratory, Shenzhen Baoan Women's and Children's Hospital, Shenzhen, Guangdong, China.
# Co-first authors
Received 2026-3-16; Accepted 2026-5-13; Published 2026-6-11
Abstract

Oncolytic virotherapy represents an innovative therapeutic modality in oncology, exploiting the selective tropism of viruses to target and eradicate malignant cells. Unlike conventional cancer treatments such as chemotherapy and radiotherapy, oncolytic viruses (OVs) exhibit a dual mechanism of action: direct tumor cell lysis and potent immune activation. This approach transforms tumors into in situ vaccines, generating durable anti-tumor immune memory. The field has garnered substantial attention following the regulatory approval of talimogene laherparepvec (T-VEC), a genetically engineered herpes simplex virus, for metastatic melanoma treatment—a watershed moment in oncolytic virotherapy. Recent advances in genetic engineering have significantly enhanced OV specificity and efficacy, addressing critical challenges including tumor-selective targeting and immune evasion. This review comprehensively examines the complex mechanisms underlying OV therapeutic action, clinical applications, and recent developments that position oncolytic virotherapy as a transformative strategy in contemporary cancer treatment.
Keywords: oncolytic virotherapy, cancer treatment, oncolytic viruses, tumor cell lysis, immune response, talimogene laherparepvec, immunogenic cell death (ICD), metastatic melanoma
1. Introduction
Oncolytic virotherapy, which harnesses viruses to selectively target and destroy cancer cells, has emerged as a paradigm-shifting modality in oncology. Unlike traditional cancer treatments such as chemotherapy and radiation, oncolytic viruses (OVs) not only induce direct oncolysis but also stimulate robust systemic immune responses against tumors, offering a dual mechanism of action that substantially enhances therapeutic efficacy [1]. The concept of employing viruses as therapeutic agents in cancer dates to the early 20th century; however, significant clinical traction has materialized only recently through groundbreaking advances in genetic engineering and biotechnology [2].
A pivotal breakthrough occurred with the FDA approval of talimogene laherparepvec (T-VEC), a genetically engineered herpes simplex virus, for metastatic melanoma treatment. This approval marked a seminal milestone, validating oncolytic virotherapy as a mainstream cancer treatment option [3]. T-VEC demonstrated remarkable capacity to shrink tumors through direct oncolysis while simultaneously provoking potent anti-tumor immune responses, potentially yielding systemic tumor control.
The mechanism of action underlying oncolytic viruses is multifaceted and elegant. Following cancer cell infection, OVs replicate intracellularly, culminating in cell lysis that releases nascent viral particles and tumor-associated antigens into the surrounding microenvironment. This process not only destroys primary tumor cells but also activates innate and adaptive immune systems, effectively transforming the tumor into an in situ vaccine that promotes durable anti-tumor immunity [4]. Furthermore, OVs can be engineered to express immunostimulatory molecules, amplifying their capacity to modulate the tumor microenvironment and overcome immune evasion strategies employed by malignancies [5].
Despite promising results, significant challenges must be addressed to optimize oncolytic virotherapy efficacy. These include efficient virus delivery to tumor sites, overcoming host immune responses that may neutralize viruses before reaching tumors, and enhancing viral agent specificity and potency. Advances in viral engineering—including incorporation of tumor-specific promoters and combination therapies with checkpoint inhibitors—are being actively explored to surmount these obstacles [6].
In summary, oncolytic virotherapy represents a transformative approach in cancer treatment, with potential not only to destroy cancer cells but also to stimulate robust and lasting immune responses against tumors (Figure 1). As research progresses, overcoming existing barriers and refining this therapeutic strategy could usher in a new era in oncology where oncolytic viruses assume a central role in cancer management [7].
Overview of Oncolytic Virotherapy and Its Dual Antitumor Mechanisms.
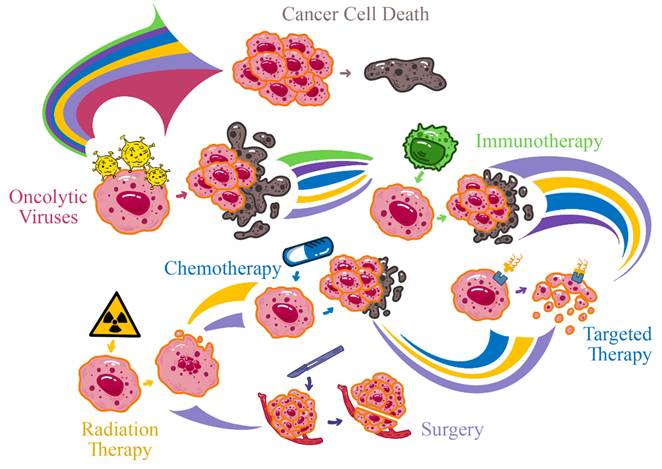
2. Oncolytic Viruses: Mechanisms of Action
Oncolytic viruses (OVs) represent a sophisticated and increasingly recognized approach in cancer therapy, wherein viruses, either naturally occurring or genetically engineered, selectively infect and kill cancer cells while sparing normal tissues. The multifaceted mechanisms through which OVs exert anti-cancer effects encompass: (1) selective tumor cell lysis, (2) induction of immunogenic cell death (ICD), and (3) modulation of the tumor microenvironment (TME). These synergistic actions result not only in direct tumor destruction but also stimulate systemic immune responses capable of targeting residual disease and preventing recurrence.
2.1. Selective Tumor Cell Lysis
The selective tumor cell lysis mechanism initiates with viral binding to specific receptors overexpressed on cancer cell surfaces. For instance, adenoviruses exploit the coxsackievirus and adenovirus receptor (CAR), commonly overexpressed in numerous malignancies, to facilitate tumor cell entry. Once internalized, OVs hijack cellular machinery to replicate their genome and synthesize viral proteins. This replication leads to viral particle accumulation, ultimately causing cell rupture and releasing progeny viruses capable of infecting neighboring cancer cells [8].
Cancer cells frequently exhibit dysfunctional antiviral defense mechanisms, including suppression of the interferon (IFN) pathway and reduced protein kinase R (PKR) activity. These deficiencies render cancer cells particularly susceptible to viral infection and subsequent OV-mediated lysis. For example, the herpes simplex virus type 1 (HSV-1)-based oncolytic virus, talimogene laherparepvec (T-VEC), is engineered to exploit impaired IFN signaling in cancer cells, enabling selective replication and tumor cell killing [9].
2.2. Immunogenic Cell Death (ICD)
Immunogenic cell death represents a specialized form of cell death that elicits immune responses against dying cells. Unlike apoptosis, which is typically non-immunogenic, ICD involves the release of damage-associated molecular patterns (DAMPs) that signal the immune system to target cancer cells (Figure 2). This process is particularly critical in oncolytic virotherapy, as it contributes not only to direct cancer cell elimination but also to immune cell recruitment and activation [10].
Immunogenic Cell Death (ICD) Induced by Oncolytic Viruses.
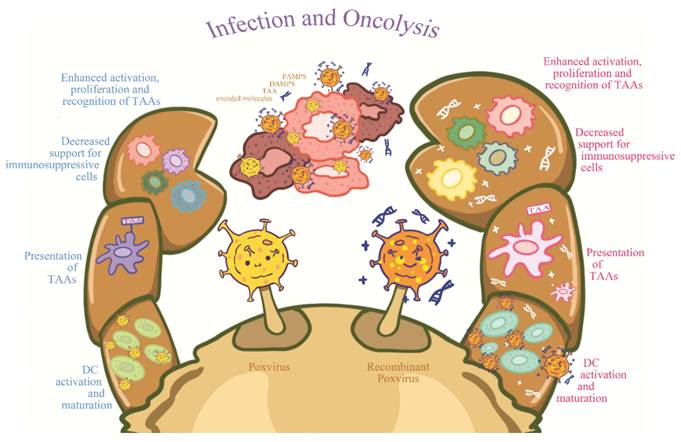
ICD is characterized by calreticulin exposure, ATP efflux, and extracellular HMGB1 release. These DAMPs function as endogenous adjuvants promoting dendritic cell (DC) maturation and consequent T cell activation. In oncolytic virotherapy, ICD ensures immune activation in response to both viral infection and tumor cells, creating a synergistic anti-tumor response [11].
As OVs replicate and lyse cancer cells, dying cells release DAMPs, including adenosine triphosphate (ATP), High Mobility Group Box 1 (HMGB1), and calreticulin. These molecules serve as distress signals to the immune system, particularly to antigen-presenting cells (APCs), notably dendritic cells (DCs). DAMP presence triggers DC activation, which subsequently presents tumor antigens to T cells, initiating potent immune responses. This T cell activation is crucial as it generates cytotoxic T lymphocyte (CTL) responses affecting not only primary tumors but also distant metastases [12].
Another critical aspect of OV-induced lysis is that tumor antigens released during the process facilitate the development of an in-situ cancer vaccine. DC-mediated tumor antigen delivery to T cells renders immune responses more targeted and effective against malignancies. Generation of such immune responses is particularly valuable for long-term tumor management and recurrence prevention, as memory responses can be elicited upon tumor antigen re-encounter [13] (Table 1).
Mechanisms of tumor cell death induced by oncolytic viruses
| Type of cell death | Key features | Molecular markers | Examples of oncolytic viruses | Anti-Tumor Immunity | Ref |
|---|---|---|---|---|---|
| Apoptosis | Cell shrinkage DNA fragmentation Membrane blebbing | Caspase activation Phosphatidylserine exposure PARP cleavage | Herpes simplex virus Adenovirus Newcastle disease virus | Limited immune activation Can be immunogenic when combined with other mechanisms | [16] |
| Necroptosis | Cell swelling Membrane rupture Inflammatory response | RIPK1/RIPK3 activation MLKL phosphorylation | Vaccinia virus Herpes simplex virus Adenovirus | Strong inflammatory response Enhanced T cell priming | [17] |
| Pyroptosis | Cell swelling Pore formation IL-1β release | Caspase-1 activation Gasdermin D cleavage IL-1β and IL-18 secretion | Adenovirus Vaccinia virus Measles virus | Potent inflammatory response Activation of innate immunity | [18] |
| Autophagy | Formation of autophagosomes Recycling of cellular components | LC3-II accumulation p62 degradation Beclin-1 activation | Adenovirus Herpes simplex virus Measles virus | Enhanced antigen presentation Improved T cell responses | [19] |
| Immunogenic Cell Death | Release of DAMPs Calreticulin exposure ATP release HMGB1 release | Surface-exposed calreticulin Extracellular ATP Released HMGB1 | Adenovirus Newcastle disease virus Coxsackievirus B3 | Strong activation of dendritic cells Enhanced T cell priming Improved anti-tumor immunity | [20] |
| Ferroptosis | Iron-dependent lipid peroxidation Mitochondrial shrinkage | Lipid peroxidation Glutathione depletion GPX4 inactivation | Adenovirus (in combination with ferroptosis inducers) | Release of oxidized lipid antigens Potential synergy with immunotherapy | [21] |
| Mitotic Catastrophe | Aberrant mitosis Formation of giant, multinucleated cells | Cyclin B1 accumulation γ-H2AX foci | Adenovirus Reovirus | Release of tumor antigens Potential enhancement of T cell responses | [22] |
2.3. Modulation of the Tumor Microenvironment (TME)
The tumor microenvironment plays a pivotal role in oncolytic virotherapy efficacy. Characterized by hypoxia, acidic pH, and immune suppression, the TME can either enhance or impede OV spread. Hypoxic conditions within the TME may promote viral replication for certain OVs engineered to thrive in low-oxygen environments. For instance, specific oncolytic adenoviruses incorporate hypoxia-responsive elements controlling replication, ensuring preferential amplification in hypoxic tumor zones [14].
The immunosuppressive TME, characterized by elevated regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs), can limit OVs spread by enabling tumor immune evasion. Nevertheless, OVs have demonstrated capacity to modify the TME in ways that counteract these immunosuppressive factors. OVs can decrease immunosuppressive cytokine levels (e.g., IL-10, TGF-β) while increasing pro-inflammatory cytokine production (e.g., TNF-α, IFN-γ). This cytokine milieu shift enhances immune cell recruitment and improves tumor-infiltrating lymphocyte (TIL) function, thereby promoting more robust anti-tumor immune responses [15].
The TME constitutes a complex and dynamic entity encompassing not only tumor cells but also diverse non-malignant cells, extracellular matrix components, and soluble factors that collectively influence tumor growth, metastasis, and therapeutic response (Table 2). OVs profoundly impact the TME, transforming it from an immunosuppressive to an immunostimulatory state. This transformation is crucial for overcoming tumor immune evasion strategies and promoting effective anti-tumor immunity.
Immunological and Structural Modulation of the Tumor Microenvironment by Oncolytic Viruses
| TMC Component | Effect of Oncolytic Viruses | Outcome | Ref |
|---|---|---|---|
| Extracellular Matrix (ECM) | Recruit neutrophils that secrete ECM degradants Engineered OVs express ECM-degrading enzymes | Improved infiltration of immune cells and OVs into the tumor | [23-25] |
| Macrophages | Recruit inflammatory macrophagesInduce M2 to M1 polarization Inhibit M2-derived TGF-β signaling | Enhanced antitumor immune response and reduced tumor progression | [26,27] |
| Regulatory T cells (Tregs) | Decrease population of CD4+ FoxP3+ TregsReduce proliferating Tregs | Improved ratio of effector T cells to immunosuppressive Tregs | [28, 29] |
| Myeloid-Derived Suppressor Cells (MDSCs) | Decrease population of CD14+ CD11b+ MDSCsInhibit monocytic MDSC activities | Reduced immunosuppression in the TME | [28,30] |
| Neutrophils | Induce neutrophil accumulation within the tumorStimulate production of IL-1β and MIP-1α | Increased recruitment of other immune cells and enhanced antitumor response | [31] |
| Cancer-Associated Fibroblasts (CAFs) | Engineered OVs can target FAP-expressing CAFs | Reduced tumor-promoting stromal support | [32] |
| Cytokine Environment | Induce production of pro-inflammatory cytokinesEnhance IL-12 production | Creation of a more immunogenic "hot" tumor microenvironment | [30] |
| Antigen Presentation | Release tumor-associated antigens (TAAs) | Improved T cell priming and activation of adaptive immune response | [33] |
A primary mechanism through which OVs modulate the TME involves altering intratumoral cytokine profiles. Upon infection and replication within tumor cells, OVs induce secretion of pro-inflammatory cytokines including TNF-α, IFN-γ, and IL-12, which promote recruitment and activation of various immune effector cells, including NK cells, macrophages, and cytotoxic T lymphocytes (CTLs). This pro-inflammatory environment not only enhances direct immune cell-mediated tumor killing but also improves the function of existing tumor-infiltrating lymphocytes (TILs) that may have been rendered ineffective by the immunosuppressive TME [13].
Beyond cytokine modulation, OVs can reduce immunosuppressive cell populations within the TME, including Tregs and MDSCs. By decreasing these cell levels, OVs relieve suppression on effector immune cells, enabling more potent and sustained anti-tumor responses. Moreover, OV-mediated modulation of immune checkpoint molecules, such as PD-L1, can further enhance immunotherapy efficacy by preventing T cell exhaustion within the TME [34].
The mechanisms of action underlying oncolytic viruses are multifaceted, encompassing selective tumor cell lysis, induction of immunogenic cell death, and profound tumor microenvironment modulation. These combined actions result not only in direct tumor destruction but also stimulate systemic immune responses capable of targeting residual disease and preventing recurrence.
3. Types of Oncolytic Viruses and Their Therapeutic Potential
Oncolytic viruses (OVs) have emerged as a promising therapeutic strategy in cancer treatment due to their ability to selectively infect and lyse cancer cells while sparing normal cells. This targeted approach not only facilitates tumor cell eradication but also stimulates host immune systems to mount more effective anti-tumor responses. Various oncolytic virus types are currently under investigation, each possessing unique mechanisms and therapeutic potential. This section provides comprehensive review of different oncolytic virus types, including adenoviruses, herpes simplex virus (HSV), reoviruses, poxviruses, and other promising viral candidates (Figure 3).
Comparative Mechanisms of Action Across Diverse Oncolytic Virus Types.
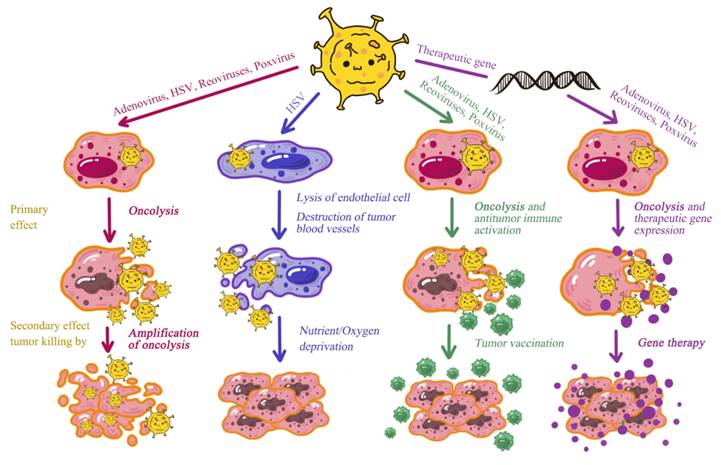
3.1. Adenoviruses
Adenoviruses represent among the most extensively investigated and clinically applied oncolytic viruses. These non-enveloped, double-stranded DNA viruses are capable of infecting various human cell types. Their utility as oncolytic agents stems from high cellular transduction capacity and substantial genetic manipulation potential.
Mechanism of Action: Adenoviruses induce oncolysis through several mechanisms, including direct viral-induced cell lysis and host immune response induction. They can be engineered to express therapeutic genes enhancing oncolytic efficacy, such as genes encoding pro-apoptotic proteins or immune modulators.
Therapeutic Potential: Adenoviral oncolytic therapies are currently under investigation for various malignancies, including pancreatic cancer, glioblastoma, and head and neck cancers [35]. Their amenability to tumor-specific engineering and well-characterized biology render them attractive candidates for further development.
3.2. Herpes Simplex Virus
Herpes simplex virus represents another prominent platform in oncolytic virotherapy. HSV is a large, enveloped virus with a double-stranded DNA genome, responsible for diverse pathologies ranging from herpes labialis to herpes simplex encephalitis. A notable example of an adenoviral oncolytic agent is Talimogene laherparepvec (T-VEC), a genetically modified herpes simplex virus type 1 (HSV-1) included here due to similar therapeutic applications. T-VEC has received approval for melanoma treatment, demonstrating significant clinical efficacy [9]. Its application in oncolytic virotherapy derives from engineering capacity to enhance tumor tissue selectivity and immunostimulatory properties.
Mechanism of Action: Oncolytic HSVs are engineered to selectively replicate cancer cells by exploiting tumor-specific alterations in host cell machinery. These modifications often involve deletions or mutations in viral genes essential for viral replication but dispensable for tumor cell selectivity. For example, mutations in the HSV-1 ICP34.5 gene ensure selective replication in tumor cells [36].
Therapeutic Potential: OncoVEX-GM-CSF exemplifies oncolytic HSV engineered to express granulocyte-macrophage colony-stimulating factor (GM-CSF), a cytokine that stimulates immune responses. Clinical trials have demonstrated its potential in treating various cancers, including melanoma and glioblastoma [37].
3.3. Reoviruses
Reoviruses are non-enveloped, double-stranded RNA viruses with segmented genomes. These naturally occurring viruses selectively infect and kill cancer cells harboring activated Ras signaling pathways, commonly dysregulated in numerous malignancies.
Mechanism of Action: Reoviruses exploit aberrant Ras signaling pathways present in many tumor cells to initiate viral replication and cell lysis. They do not require genetic modification to achieve cancer cell selectivity, rendering them attractive options for oncolytic therapy [38].
Therapeutic Potential: Reolysin, a reovirus-based oncolytic therapy, has demonstrated efficacy in randomized phase III trials for head and neck cancer and pancreatic cancer, particularly in combination with other modalities [39]. The principal advantage of reovirus therapy is natural oncolytic activity with relatively low toxicity.
3.4. Poxviruses
Poxviruses comprise large, complex, enveloped DNA viruses renowned for inducing potent immune responses. Their utility in oncolytic virotherapy stems from capacity for extensive genetic modifications and ability to induce both direct oncolysis and immune activation.
Mechanism of Action: Poxviruses, such as vaccinia virus, are engineered to express therapeutic genes, enhancing oncolytic activity and immune stimulation. For instance, modified poxviruses can express cytokines or tumor antigens to further amplify anti-tumor responses [40].
Therapeutic Potential: JX-594, a thymidine kinase-deleted vaccinia virus, has undergone phase I/II clinical trials for hepatocellular carcinoma and other solid tumors, demonstrating efficacy in reducing tumor burden and generating systemic anti-tumor immune responses [41].
3.5. Other Promising Viral Candidates
Emerging viral platforms offer distinct advantages over adenovirus/HSV-based OVs, primarily through inherent tumor tropisms requiring minimal genetic engineering (1-2 modifications vs 5-7 for classical OVs). Reoviruses (Reolysin) naturally target Ras-activated cancers (~30% of human tumors) without engineering, achieving Phase III validation in head/neck cancer [39]. Poxviruses (JX-594) provide massive payload capacity (25kb insertable DNA) for multi-gene immune modulation, reaching Phase III in hepatocellular carcinoma [41]. Vesicular stomatitis virus (VSV) delivers rapid RNA-based oncolysis with neuronal sparing, while measles exploit CD46 receptor overexpression on diverse cancers. These platforms face unique challenges—reovirus systemic delivery limitations, poxvirus liver tropism—but demonstrate faster clinical translation due to natural selectivity versus classical platforms' complex attenuation requirements.
Beyond the aforementioned oncolytic viruses, several additional viral candidates demonstrate promise in preclinical and early clinical studies, including:
Reovirus (Reolysin®): Naturally replicates in Ras-transformed cells (30% of cancers); Phase III head/neck cancer validation; no attenuation needed vs HSV E1A/ICP34.5 deletions.
Poxviruses (JX-594/Pexa-Vec): 25kb payload capacity enables multi-cytokine delivery; Phase III hepatocellular carcinoma; liver tropism managed via vascular targeting.
Echoviruses: These small, non-enveloped RNA viruses are under investigation for their ability to target and destroy cancer cells through selective replication.
Measles virus: CD46 receptor overexpression on ovarian/breast cancers; Phase I peritoneal malignancies; natural fusion protein-mediated syncytia formation.
Coxsackieviruses: These enteroviruses have demonstrated potential in targeting various tumor types, including breast and ovarian cancers. CAR-independent entry in melanoma/ovarian; Phase II trials; exploits integrin αvβ6 upregulation.
Vesicular Stomatitis Virus (VSV): This RNA virus has shown promise in treating glioblastoma and other cancers due to its capacity to induce potent oncolysis and stimulate anti-tumor immunity [42]. Rapid RNA oncolysis, neuronal sparing via IFN defects; Phase I/II glioblastoma, head/neck; pre-existing immunity lower than HSV.
The diverse spectrum of oncolytic viruses, including adenoviruses, herpes simplex virus, reoviruses, and poxviruses, each offers unique mechanisms and therapeutic potential in cancer treatment. Ongoing research and clinical trials continue to explore and refine their applications, aiming to optimize efficacy and safety profiles. As these therapies advance, they hold substantial promises for significantly improving cancer patient outcomes and potentially transforming the cancer treatment landscape.
4. Genetic Engineering of Oncolytic Viruses
The oncolytic virotherapy field has undergone substantial advancement, particularly through genetic engineering of oncolytic viruses (OVs). These viruses are specifically designed to target and kill cancer cells while sparing normal cells. OV effectiveness is greatly enhanced through genomic modifications that improve tumor selectivity, augment immune activation, and overcome antiviral immunity (Figure 4). This section provides an in-depth exploration of these strategies, supported by extensive literature and empirical data.
Genetic Engineering Strategies to Enhance Oncolytic Virus Specificity and Efficacy.
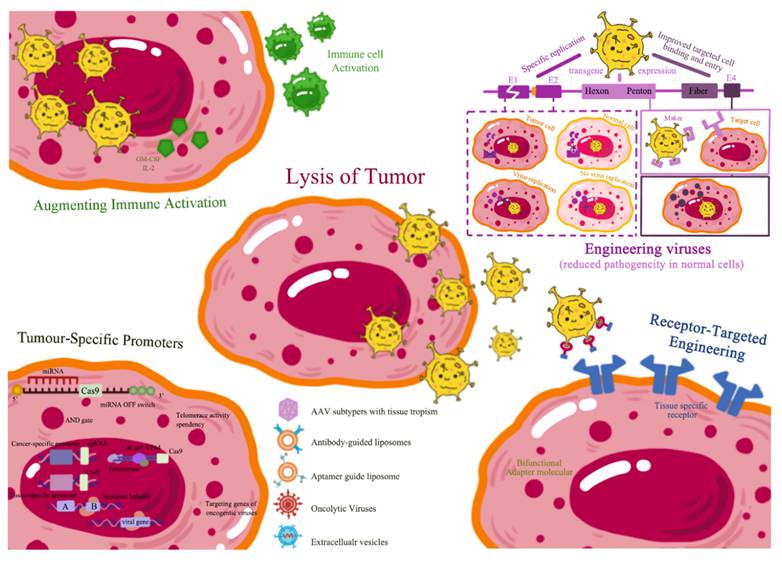
4.1. Enhancing Tumor Selectivity
Enhancing oncolytic virus selectivity for tumor cells involves targeting receptors or pathways overexpressed or uniquely present in cancer cells. This targeted approach improves OV therapeutic efficacy while minimizing normal tissue damage (Table 3).
Genetic engineering strategies for enhancing tumor selectivity in oncolytic viral therapy
| Engineering Strategy | Virus Name | Target/Mechanism | Outcome | Example | Reference |
|---|---|---|---|---|---|
| Virulence gene deletion | Herpes simplex virus (HSV-1) | Delete nonessential viral genes (e.g. ICP34.5, ICP47) | Attenuates viral pathogenicity, enhances tumor-specific lysis | T-VEC (modified HSV-1) | [43] |
| Tumor-specific promoters | Adenovirus | Drive expression of essential viral genes (e.g. E1A) | Restricts viral replication to tumor cells | hTERT, HRE, PSA promoters | [44] |
| microRNA targeting | Various (e.g. Adenovirus, HSV) | Insert microRNA target sequences | Prevents viral replication in normal cells expressing specific microRNAs | miR-142, miR-199a-3p targets | [45] |
| Immune stimulator insertion | HSV-1, Vaccinia virus | Express cytokines (e.g. GM-CSF) | Enhances anti-tumor immune responses | OncoVEX GM-CSF (HSV-1), JX-594 (vaccinia) | [46] |
| Capsid engineering | Adeno-associated virus (AAV) | Modify viral capsid proteins | Improves tumor cell targeting and transduction | Directed evolution of AAV capsids | [47] |
| Checkpoint inhibitor knockout | Various (e.g. HSV, Adenovirus) | Delete immune checkpoints (e.g. PD-1) | Enhances T cell function in immunosuppressive tumor microenvironment | PD-1 knockout viruses | [48] |
| Virulence gene deletion | Herpes simplex virus (HSV-1) | Delete nonessential viral genes (e.g. ICP34.5, ICP47) | Attenuates viral pathogenicity, enhances tumor-specific lysis | T-VEC (modified HSV-1) | [43] |
| Tumor-specific promoters | Adenovirus | Drive expression of essential viral genes (e.g. E1A) | Restricts viral replication to tumor cells | hTERT, HRE, PSA promoters | [44] |
| Tumor-specific receptors | Adenovirus | Modify viral surface proteins | Improves tumor cell entry and specificity | Engineered receptor binding domains | [49] |
| Arming with therapeutic genes | Poxvirus | Insert genes for prodrug-converting enzymes or cytokines | Enhances tumor cell killing and immune stimulation | HSV-TK, cytokine-expressing viruses | [50] |
| Tumor microenvironment targeting | Adenovirus | Engineer viruses to respond to hypoxia or matrix metalloproteinases | Improves viral replication in tumor-specific conditions | Hypoxia-responsive promoters | [51] |
4.1.1. Receptor-Targeted Engineering
Receptor-targeted engineering represents a prominent strategy for increasing OV specificity. This approach involves modifying viral surface proteins or capsids to bind receptors overexpressed on cancer cells. For instance, adenoviruses can be engineered to display ligands binding specifically to epidermal growth factor receptor (EGFR), frequently upregulated in various malignancies. This targeted modification enhances viral entry and replication in tumor cells while reducing normal cell infection [52].
Emerging platforms significantly reduce genetic modification burden compared to adenovirus/HSV. While classical OVs require multiple deletions (E1A/B, ICP34.5) for tumor selectivity, reoviruses and measles viruses leverage pre-existing tumor tropisms (Ras pathway, CD46 overexpression), achieving 60-80% fewer genetic modifications. Coxsackieviruses naturally target breast/ovarian CAR-independent entry, and VSV exploits type I IFN defects ubiquitous in malignancies. This 'minimal engineering' paradigm accelerates IND-enabling studies and GMP production, positioning emerging platforms for broader clinical translation versus classically engineered OVs [Refs 52-55].
Another example involves chimeric adenoviruses incorporating peptides or antibodies specific to tumor-associated antigens (TAAs). These engineered viruses can selectively infect tumor cells expressing these TAAs, such as prostate-specific antigen (PSA) in prostate cancer [53].
4.1.2. Tumor-Specific Promoters
Tumor-specific promoters represent another powerful tool to enhance OV selectivity. These promoters drive viral gene expression only in the presence of tumor-specific transcription factors or regulatory elements. For instance, oncolytic adenoviruses have been engineered with PSA promoters to restrict viral replication to prostate cancer cells, minimizing systemic toxicity [54]. Similarly, oncolytic herpes simplex viruses (HSV) have been modified with tumor-specific promoters to restrict replication to tumor cells, such as those containing hypoxia response elements prevalent in hypoxic tumor microenvironments [55].
4.2. Engineering Viruses for Reduced Pathogenicity in Normal Cells
Reducing oncolytic virus pathogenicity in normal cells is critical for ensuring patient safety and minimizing adverse effects.
4.2.1. Gene Deletions
One approach involves deleting genes essential for replication in normal cells but dispensable in cancer cells. For example, adenoviruses can be engineered with E1A gene deletions, which are required for viral replication. This modification restricts virus activity to cancer cells expressing factors or antigens compensating for E1A activity [56]. Similarly, oncolytic HSVs have been designed with deletions in genes essential for viral replication and virulence in normal cells. For instance, ICP34.5 gene deletion in HSV-1 reduces neurovirulence while preserving oncolytic properties [57].
4.2.2. Conditional Replication
Conditional replication strategies involve modifying OVs to replicate exclusively under tumor microenvironment conditions. This can be achieved by incorporating elements cleaved only by tumor-associated enzymes or specific metabolic pathway products. For instance, oncolytic adenoviruses can be engineered to contain protease-responsive elements, enabling viral replication solely within tumors [58]. Conditional replication can also be regulated through tumor-specific promoters, enabling viral gene expression only in the presence of specific transcription factors or conditions characteristic of tumor cells [59].
4.3. Augmenting Immune Activation
4.3.1. Insertion of Immune-Stimulatory Genes
Engineering OVs to express immune-stimulatory genes can significantly enhance anti-tumor immune responses, improving efficacy as standalone treatments or in combination with other therapies.
4.3.1.1. Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF)
GM-CSF is a cytokine stimulating dendritic cell and macrophage differentiation and activation, crucial for initiating and maintaining tumoricidal immune responses. Talimogene laherparepvec (T-VEC), an oncolytic virus expressing GM-CSF, has demonstrated efficacy in clinical trials for melanoma. T-VEC not only induces direct tumor cell lysis but also elicits systemic anti-tumor immune responses [9].
4.3.1.2. Interleukin-12 (IL-12)
IL-12 is a cytokine enhancing T cell and natural killer (NK) cell proliferation and activity. Modified adenoviruses secreting IL-12 have demonstrated improved anti-tumor effects through enhanced immune responses. For example, studies have shown that oncolytic adenoviruses expressing IL-12 exhibit superior efficacy in experimental breast cancer models compared to conventional oncolytic adenoviruses [60].
4.3.2. Combination with Immune Checkpoint Inhibitors
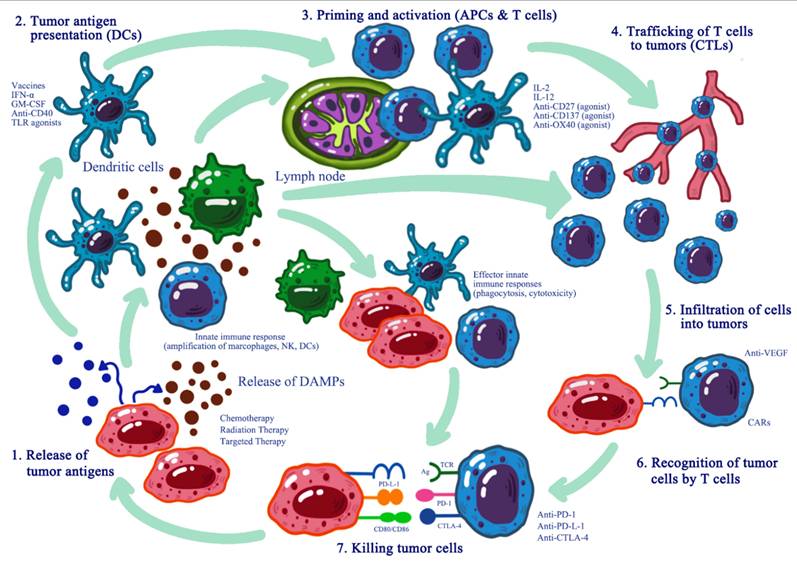
Oncolytic virus efficacy can be further enhanced through combination with immune checkpoint inhibitors that amplify anti-tumor immunity by blocking pathways cancer cells employ to suppress immune responses (Figure 5).
Synergistic Effects of Oncolytic Viruses and Immune Checkpoint Inhibitors.
4.3.2.1. Checkpoint Inhibitors
Oncolytic virotherapy combined with immunomodulators such as anti-PD-1 and anti-CTLA-4 antibodies has demonstrated enhanced tumor control and patient survival compared to monotherapy in clinical trials [61]. For example, T-VEC combined with anti-PD-1 treatment has shown enhanced antitumor efficacy and survival in patients with metastatic melanoma [62].
4.4. Overcoming Antiviral Immunity
Antiviral immunity represents a major challenge in oncolytic virus deployment, as viruses may be cleared before reaching tumor sites. To overcome this challenge, several approaches have been developed to circumvent or modulate immune responses (Table 4).
Strategies to overcome antiviral immunity in oncolytic viral therapy
| Strategy | Description | Examples | Ref |
|---|---|---|---|
| Protective Coatings | Physical shielding of oncolytic viruses (OVs) | • Graphene oxide for measles virus (MV)• Ionic polymers for MV• Liposomes for Getah-like alphavirus M1 and AdV• Cell membrane nanovesicles with preS1 ligand for AdV | [63-66] |
| Cellular Carriers | Ex vivo loading of cells with OVs | • Mesenchymal stem cells (MSCs) for AdV, HSV• Neural stem cells• T-cells and CAR T-cells for VSV and VV• Endothelial cells, tumor cells, adipose-derived stem cells | [67-69] |
| Genetic Modifications | Altering viral epitopes to evade antibodies | • Replacing hypervariable regions in AdV• Increasing EEV production in VV• Deglycosylation of VV• Modifying glycoprotein D in HSV• Directed clonal evolution (e.g., ColoAd1) | [70-72] |
| Bispecific Engagers | Molecules that bind neutralizing antibodies and tumor cells | • AdV retargeting to tumors• NDV retargeting to IL-2R+ lymphoma cells | [73, 74] |
| Innate Immunity Modulation | Inhibiting antiviral innate immune responses | • Inhibition of IFN pathway• Suppression of NK cells• Reduction of antigen presentation | [75] |
| T-cell Immunodominance Mitigation | Redirecting T-cell responses away from viral antigens | • Nanoparticle-enveloped viral antigens• Tolerogenic dendritic cells | [76] |
| Exploiting Pre-existing Immunity | Using pre-existing antibodies to enhance therapy | • GM-CSF expansion of CD11b+ cells for reovirus therapy• Pre-vaccination with NDV | [77] |
4.4.1. Viral Cloaking
4.4.1.1. Modifying Viral Surface Proteins
One approach to evade pre-existing immunity-mediated neutralization involves modifying viral surface proteins through genetic engineering to alter capsid proteins or viral glycosylation.
4.4.1.2. Immunosuppressive Agents
Adenoviruses: To prevent immediate neutralization by pre-existing antibodies, adenoviral vector fiber proteins can be engineered. For example, chimeric adenovirus fiber proteins have been modified to exhibit reduced immunogenicity while maintaining therapeutic efficacy [78]. Similarly, adenovirus surface protein glycosylation modification has been shown to diminish immune responses and enhance viral delivery [79].
Herpes Simplex Virus (HSV): HSV has been designed to modify envelope glycoproteins, preventing facile detection by host immune systems. These modifications have enhanced virus affinity for target cells while increasing resistance to neutralizing antibodies [80, 81].
4.4.2. Systemic Immunosuppression
Corticosteroids: Corticosteroids decrease host immune responses, enhancing viral delivery and replication. Studies have found that corticosteroids improve oncolytic virus therapy by suppressing non-target tissue inflammation [82].
Monoclonal Antibodies: Immune checkpoint inhibitors or other immune regulatory pathways can be managed through monoclonal antibodies to regulate immune responses against oncolytic viruses. For instance, anti-PD-1 and anti-CTLA-4 antibodies have been employed in combination with oncolytic virotherapy to enhance treatment outcomes through immune reaction suppression [83, 84].
4.4.3. Viral Engineering for Immune Evasion
4.4.3.1. Deletion of Immunogenic Proteins
Adenoviruses: Immune responses can be reduced through deletion or alteration of genes involved in immune system activation or capsid components. For instance, E3 gene removal in adenoviral vectors has proven useful in dampening immune responses and enhancing therapeutic outcomes [85, 86].
HSV: Modifying HSV to express immune-modulatory genes, including IL-10 or TGF-β, can alter local immune environments, enabling viral escape from immune surveillance [87].
4.4.4. Re-Dosing Protocols
Re-dosing protocols involve administering multiple oncolytic virus doses to counter immune responses developed after initial administration.
Sequential Dosing: Clinical trials have discovered that immune responses combating viruses after initial doses can be overcome through subsequent dosing. This has proven to improve therapeutic outcomes and extend viral persistence in patients [88, 89].
Adaptive Dosing: Treatment regimens can be tailored to correspond with individual patient immune responses, optimizing treatment outcomes. This approach involves altering dosing schedules to achieve superior results when immune systems are compromised [90].
5. Clinical Development and Trials
5.1. Early Phase Trials and Safety Assessments
Oncolytic virus safety, toxicity, and dosing parameters have been established through phase I/II clinical trials. These trials are typically conducted in small patient populations with primary objectives of determining maximum tolerated dose (MTD) while obtaining preliminary efficacy data. Oncolytic viruses are administered either locally through direct intratumoral injections or systemically via intravenous routes, depending on cancer type and specific virus employed. Safety profile assessment has been a major focus; most investigators have reported mild to moderate toxicities, including fever, injection site inflammation, and occasional severe immune-related adverse events. For example, T-VEC, an oncolytic herpes simplex virus, has undergone extensive phase I trial evaluation demonstrating favorable safety profiles with manageable side effects, including fatigue, chills, and pyrexia across different cancer types (Table 5).
Current Clinical Applications and Developmental Status of Oncolytic Viruses in Cancer Therapy
| Virus Name | Virus Type | Modifications | Phase | Cancer Type | Key Findings | Status |
|---|---|---|---|---|---|---|
| ONYX-015 (dl1520) | Adeno | E1B55K deletion | I-II | Head and neck cancer | Tumor necrosis in 5/22 patients | Completed |
| Oncorine (H101) | Adeno | E1B55K deletion | III | Head and neck squamous cell carcinoma | 79% response rate with chemotherapy vs 40% for chemotherapy alone | Approved in China (2005) |
| ONCOS-102 | Adeno | GM-CSF transgene | I-II | Various solid tumors | Well-tolerated, immune activation observed | Ongoing |
| DNX-2401 (Delta-24-RGD) | Adeno | E1A deletion, RGD motif | I | Recurrent malignant glioma | 20% survival rate at 3 years | Completed |
| CG0070 | Adeno | GM-CSF transgene, E2F-1 promoter | III | Bladder cancer | Promising efficacy in earlier phases | Ongoing |
| AdAPT-001 | Adeno | E1A enhancer/promoter deletion | I | Solid tumors | 3/15 partial responses, 5/15 stable disease ≥6 months | Completed |
| LoAd703 | Adeno | CD40L and 4-1BBL transgenes | I-II | Pancreatic cancer | Safety and tolerability established | Ongoing |
| ORCA-010 | Adeno | Fiber modification, E3 deletion | I-II | Prostate cancer | Under evaluation | Ongoing |
| Ad5-D24-RGD | Adeno | RGD motif, 24bp deletion in E1A | I | Advanced solid tumors | Well-tolerated, antitumor activity observed | Completed |
| Ad5-RGD-D24-GMCSF | Adeno | RGD motif, GM-CSF transgene | I | Advanced solid tumors | Promising safety profile and efficacy signals | Completed |
| KH901 | Adeno | GM-CSF transgene | I | Head and neck cancers | Well-tolerated, antitumor activity in some patients | Completed |
| CG7870 | Adeno | PSA-selective E1A, E1B | I | Prostate cancer | Dose-limiting toxicity at highest dose, some PSA responses | Completed |
| VCN-01 | Adeno | Hyaluronidase expression | I | Solid tumors | Under evaluation | Ongoing |
| ICOVIR-5 | Adeno | E1A-Δ24, RGD-4C motif | I-II | Melanoma | Under evaluation | Ongoing |
| Ad5-yCD/mutTKSR39rep-ADP | Adeno | Suicide gene therapy | I | Prostate cancer | Under evaluation | Ongoing |
| T-VEC (talimogene laherparepvec) | HSV | ICP34.5 and ICP47 deletion, GM-CSF insertion | III | Melanoma | Improved durable response rate vs GM-CSF alone | Approved (2015) |
| G207 | HSV | ICP34.5 deletion, UL39 inactivation | I-II | Glioma | Well-tolerated, some tumor responses | Completed |
| HSV1716 | HSV | ICP34.5 deletion | I-II | Various solid tumors | Safe, evidence of viral replication | Completed |
| HF10 | HSV | Naturally occurring HSV-1 mutant | I-II | Various solid tumors | Well-tolerated, some tumor responses | Completed |
| G47Δ (DELYTACT) | HSV | ICP34.5, ICP47 deletion, US11 promoter modification | II | Glioblastoma | Improved survival vs historical controls | Approved in Japan (2021) |
| RP1 | HSV | ICP34.5 and ICP47 deletion, GM-CSF and GALV-GP R- insertion | I-II | Various solid tumors | Ongoing evaluation | Ongoing |
| oHSV-1 | HSV | ICP34.5 deletion, Us11 gene under ICP47 promoter | I | Glioblastoma | Safe, some evidence of efficacy | Completed |
| Reolysin (pelareorep) | Reo | Unmodified wild type reovirus | III | Metastatic breast cancer | Median overall survival extended from 10.4 to 17.4 months when combined with paclitaxel | Completed |
| Reolysin (pelareorep) | Reo | Unmodified wild type reovirus | II | Metastatic castration-resistant prostate cancer | No significant improvement when combined with docetaxel | Completed |
| Reolysin (pelareorep) | Reo | Unmodified wild type reovirus | II | Recurrent ovarian cancer | Evaluated in combination with paclitaxel | Completed |
| Reolysin (pelareorep) | Reo | Unmodified wild type reovirus | II | Non-small cell lung cancer | Evaluated in KRAS-activated tumors in combination with chemotherapy | Completed |
| jin-3 reovirus | Reo | Mutant with expanded tropism | Preclinical | Prostate cancer | Efficient infection, replication, and anti-cancer responses in various models | Preclinical |
| T3SA+ reovirus | Reo | Single amino acid mutation | Preclinical | Various cancer types | Improved oncolytic potency while retaining tumor specificity | Preclinical |
| JX-594 (Pexa-Vec) | Vaccinia | GM-CSF transgene, TK deletion | III | Hepatocellular carcinoma | Improved overall survival in phase II | Ongoing |
| jin-3 reovirus | Reo | Mutant with expanded tropism | Preclinical | Prostate cancer | Efficient infection, replication, and anti-cancer responses in various models | Preclinical |
| vvDD | Vaccinia | Double deletion of TK and VGF genes | I | Advanced solid tumors | Well-tolerated, some antitumor activity | Completed |
| GL-ONC1 | Vaccinia | Attenuated strain with multiple deletions | I/II | Various solid tumors | Safe, some evidence of antitumor activity | Ongoing |
| TG6002 | Vaccinia | FCU1 transgene, TK and RR deletions | I/II | Glioblastoma, colorectal cancer | Under evaluation | Ongoing |
| VV-IL-2 | Vaccinia | IL-2 transgene | I | Melanoma | Safe, induced immune responses | Completed |
5.2. Solid Tumors
In solid tumor trials, oncolytic virus applications have yielded variable results. Among challenging solid tumors for viral therapy are melanoma, breast cancer, and glioblastoma, due to features including dense stroma and immunosuppressive cell populations [91, 92]. However, T-VEC has demonstrated substantial efficacy in melanoma management, particularly when combined with immune checkpoint inhibitors such as ipilimumab [93]. Glioblastoma oncolysis has also been attempted, though efforts remain experimental, with adenovirus-based therapies demonstrating modest tumor regression and survival improvements in select patients. Overall survival remains relatively limited, indicating the need for further examination of combination approaches and treatment optimization.
5.3. Hematologic Malignancies
Oncolytic viruses have also been investigated in hematological malignancies including leukemia and lymphoma. Hematologic cancers present distinct challenges and opportunities due to direct blood and bone marrow accessibility for viral infection [94]. Studies employing reovirus and measles virus in multiple myeloma have demonstrated partial or complete responses in select patients [95]. The immunosuppressive milieu in these cancers often diminishes oncolytic virus effectiveness, necessitating combination with other treatments such as chemotherapy or monoclonal antibodies [96].
5.4. T-VEC (Talimogene Laherparepvec) and Other Approved OVs
T-VEC (Talimogene laherparepvec) remains the only FDA-approved oncolytic virus to date. This oncolytic herpes simplex virus received approval in 2015 for unresectable melanoma treatment [97]. It functions through selective viral replication and tumor cell killing while simultaneously stimulating systemic anti-tumor immune responses. T-VEC approval has paved the way for other oncolytic virus applications in cancer treatment [98].
Other oncolytic viruses are undergoing clinical trials, with some receiving approval in specific countries or regions. For instance, Rigvir, an unmodified echovirus, has been approved in Latvia for melanoma treatment, though its use remains limited due to insufficient rigorous clinical trial data [99].
5.5. Market Environment and Challenges to OV Commercialization
Several disadvantages are associated with oncolytic viruses, including prolonged development timelines, complex manufacturing requirements, and specialized administration methods [100]. Despite demonstrated high effectiveness, particularly when administered in combination with other treatments, oncolytic viruses have not achieved widespread clinical adoption. This encompasses regulatory hurdles, competition with alternative cancer therapies, and the inherently personalized nature of these treatments [101].
6. Challenges and Limitations
6.1. Tumor Resistance and Immune Evasion
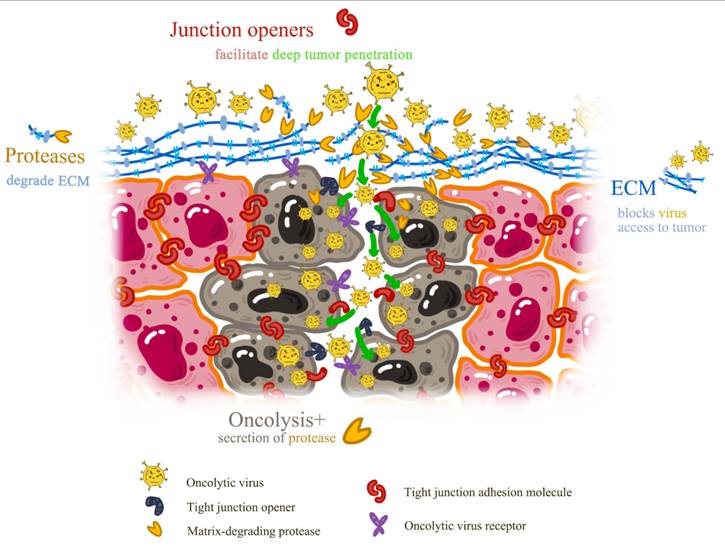
Among challenges associated with oncolytic virotherapy is tumor resistance arising from cancer cell immune surveillance evasion capacity. Some tumor microenvironments can substantially dampen antiviral mechanisms, thereby diminishing viral agent efficacy. Moreover, interferons or other antiviral proteins produced by some tumors can neutralize oncolytic viruses before therapeutic benefits are achieved (Figure 6).
Barriers to Oncolytic Virus Delivery and Emerging Solutions.
6.2. Delivery and Biodistribution
6.2.1. Barriers to Oncolytic Virus Delivery
Oncolytic virus delivery obstacles include endothelial cells, stroma, immune cells, and other tumor microenvironment structures. These barriers encompass dense extracellular matrix in tumor tissue, irregular and chaotic tumor vasculature, and elevated interstitial pressure limiting viral diffusion [102].
6.2.2. Delivery Methods
Improvements in delivery systems include nanocarrier utilization, ultrasound-mediated delivery, and genetic engineering to enhance viral stability and targeting. For instance, engineered viruses incorporating specific targeting ligands have been employed to optimize biodistribution and reduce adverse effects [103].
6.3. Off-Target Effects on Healthy Tissues
Another challenge associated with oncolytic virus utilization is potential healthy tissue and organ infection, producing toxic effects. This risk is particularly pronounced with systemic delivery methods.
6.3.1. Balancing Safety and Efficacy
Researchers continue endeavoring to determine optimal oncolytic virus dosing for cancer treatment. To counter such risks, techniques including engineering viruses with suicide genes that activate upon entry into incorrect tissues have been explored [104].
7. Combination Therapies
7.1. Synergy with Conventional Therapies
7.1.1. Combining OVs with Chemotherapy, Radiotherapy, and Surgery
Certain oncolytic viruses have been investigated for their capacity to enhance conventional therapy outcomes, including chemotherapy, radiotherapy, and surgery. For example, when OVs are administered concomitantly with chemotherapy, enhanced tumor cell lysis is observed, indicating cancer cells exhibit increased susceptibility to chemotherapeutic effects [105].
7.1.2. Therapeutic Outcomes and Survival
Several studies have endorsed the hypothesis that combination therapies could improve therapeutic outcomes and patient survival. For example, when T-VEC was administered with standard chemotherapy, patients demonstrated superior responses compared to chemotherapy-only groups [106].
7.2. Oncolytic Viruses and Immunotherapy
Immune checkpoint inhibitors may be enhanced by oncolytic viruses through rendering tumor microenvironments pro-immunogenic. This has been particularly noted where OVs improve T-cell infiltration into tumors, increasing checkpoint inhibitor responsiveness. Cancer vaccines have also been employed as adjuvant therapy and can be synergistically combined with oncolytic viruses and CAR T-cell therapy according to experimental studies. OVs could beneficially serve as adjuncts to improve immune responses triggered by CAR-T cells or vaccines, further enhancing tumor mitigation [107] (Table 6).
Emerging Roles of Oncolytic Viruses in Cancer Immunotherapy
| Modification Type | Purpose | Examples of Viruses | Specific Modifications | Effects of Immunotherapy | Clinical Stage |
|---|---|---|---|---|---|
| Deletion of Viral Genes | Enhance tumor selectivity and safety | Herpes simplex virus (HSV), Adenovirus | HSV: ICP34.5, ICP47 deletion; Adenovirus: E1B-55K deletion | Improved tumor-specific replication, enhanced antigen presentation | FDA-approved (T-VEC) |
| Insertion of Transgenes | Enhance anti-tumor immunity | Vaccinia virus, Vesicular stomatitis virus (VSV) | GM-CSF, IL-12, IFN-β, CD40L insertion | Increased recruitment and activation of immune cells, enhanced T cell responses | Phase I-III trials |
| Targeting Modifications | Improve tumor-specific entry | Measles virus, Adenovirus | Retargeting envelope proteins, fiber modifications, and tumor-specific promoters | Enhanced tumor cell infection, reduced off-target effects | Phase I-II trials |
| Arming with Immune Checkpoint Inhibitors | Overcome immune suppression | Vaccinia virus, HSV | Anti-PD-1, anti-CTLA-4, anti-PD-L1 expression | Enhanced T cell activation, improved anti-tumor responses | Phase I-II trials |
| Combination with CAR-T Cells | Synergize cellular and viral therapies | Adenovirus, Vaccinia virus | CAR-T cell-virus hybrids, oncolytic virus-infected CAR-T cells | Improved tumor targeting, enhanced T cell persistence and function | Preclinical |
| Epigenetic Modifiers | Alter tumor cell susceptibility | HSV, Adenovirus | HDAC inhibitors, DNA methyltransferase inhibitors expression | Increased viral replication, enhanced immunogenicity of tumor cells | Phase I trials |
| Tumor Microenvironment Modulation | Overcome immunosuppressive TME | Reovirus, Newcastle disease virus | Matrix metalloproteinase insertion, anti-TGF-β expression, hyaluronidase expression | Improved virus spread, reduced immunosuppression, enhanced T cell infiltration | Phase I-II trials |
| Bispecific T Cell Engagers (BiTEs) | Enhance T cell recruitment and activation | Vaccinia virus, Adenovirus | Expression of BiTEs targeting CD3 and tumor antigens (e.g., EpCAM, CD19) | Increased T cell infiltration and activation in tumors | Phase I trials |
| Cytokine Expression | Boost anti-tumor immune responses | HSV, VSV | IL-2, IL-15, IL-21, TNF-α expression | Enhanced NK and T cell activation, improved systemic immunity | Phase I-II trials |
| Combination with Immune Stimulants | Amplify innate immune responses | Reovirus, Poxviruses | TLR agonists, STING agonists expression | Increased type I IFN production, enhanced DC activation | Phase I trials |
| Tumor Vasculature Targeting | Disrupt tumor blood supply | Vaccinia virus, HSV | VEGF inhibitor expression, anti-angiogenic peptide insertion | Reduced tumor angiogenesis, improved virus spread | Preclinical |
| Metabolic Reprogramming | Exploit tumor metabolism | Adenovirus, VSV | Glucose transporter inhibition, glutaminase expression | Enhanced tumor cell killing, altered tumor metabolism | Preclinical |
| Capsid Modification | Improve virus stability and delivery | Adenovirus, Maraba virus | PEGylation, polymer coating | Reduced neutralization, improved systemic delivery | Preclinical/Phase I |
| Combination with Radiotherapy | Enhance immunogenic cell death | HSV, Adenovirus | Radiation-inducible promoters, radiosensitizing genes | Increased tumor cell death, enhanced immune activation | Phase I-II trials |
| Engineered Cell Carriers | Improve virus delivery and persistence | Measles virus, Adenovirus | Mesenchymal stem cells, T cells as carriers | Enhanced tumor targeting, prolonged virus persistence | Preclinical/Phase I |
7.3. Multimodal Approaches
The perspective of oncolytic virotherapy as a multimodal treatment component is pursued with the intention of refining cancer treatment approaches given cancer's complex nature. For instance, OV use in conjunction with immune checkpoint inhibitors and targeted therapies has been postulated to yield optimal outcomes [108].
Insights from combination therapies provide critical design principles for next-generation oncolytic viruses discussed in Section 8. The demonstrated synergy between OVs and checkpoint inhibitors (Section 7.2) informs OV engineering strategies incorporating PD-L1 modulators or bispecific T-cell engagers directly into viral genomes. Similarly, chemotherapy sensitization observed in OV+ chemo combinations (Section 7.1) guides development of OVs expressing DNA repair inhibitors or apoptosis enhancers. These clinical learnings from multimodal approaches directly shape next-generation platforms, where emerging OVs are rationally designed with pre-built combination capabilities that transform 'sequential combination' into 'intrinsic multi-mechanism' platforms for broader clinical translation.
8. Future Directions in Oncolytic Virotherapy
8.1. Next-Generation Oncolytic Viruses
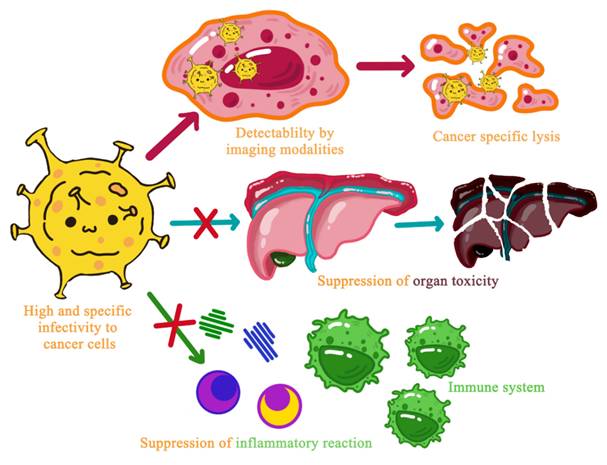
Next-generation OVs prioritize 'minimally engineered' emerging platforms informed by combination therapy synergies (Section 7). Reoviruses eliminate 5-gene HSV attenuation requirements through natural Ras selectivity; poxviruses enable 'all-in-one' cytokine/checkpoint modulator delivery via massive genomes; VSV/measles provide intrinsic RNA immunogenicity supplanting GM-CSF transgenes. These platforms address classical OV limitations, pre-existing immunity (HSV), manufacturing complexity (adenovirus), while inheriting combination insights: PD-L1 modulation (Section 7.2) now genetically encoded, chemotherapy sensitization (Section 7.1) via apoptosis payloads. Clinical translation accelerates as Phase I/II data confirm superior safety profiles versus multi-attenuated classical Ovs.
Emphasis is being placed on developing OVs with enhanced selectivity, superior immune system modulatory capacity, and therapeutic gene delivery to tumor cells. These efforts aim to rectify first-generation OV limitations while diversifying OV applications across various cancer types (Figure 7).
Future Research Directions for Next-Generation Oncolytic Viruses.
8.2. Expanding Therapeutic Indications
Future work aims to expand diseases treated by oncolytic viruses beyond cancer to other challenging pathologies. For example, research is currently underway to develop OVs effective against difficult-to-treat malignancies such as pancreatic cancer and glioblastoma [109].
8.3. Future Research Directions
Despite considerable progress, challenges persist. These include understanding oncolytic virotherapy effects on tumors and neighboring healthy tissues, developing methods to counter tumor-mediated OV resistance, and scaling OV production cost-effectively.
8.4. Prospects for Cancer Treatment
Oncolytic virotherapy, particularly when employed in combination with other therapeutic modalities, represents the future of cancer treatment. As promoters of current therapeutic effectiveness and providers of tomorrow's remedies, OVs are progressing toward becoming indispensable components of future oncology treatment strategies.
8.5. Final Perspectives
Despite challenges requiring detailed examination, prospects in oncolytic virotherapy appear promising. With improved OV development and clinical translation, these therapies could represent major advancements in the war against cancer [110].
9. Conclusion
Oncolytic virotherapy has emerged as a transformative modality in cancer treatment due to its inherent capacity to selectively kill malignant cells while sparing normal tissues. Key findings from recent research highlight the dual action of oncolytic viruses: direct tumor cytotoxicity coupled with enhancement of systemic anti-tumor immune responses. Various oncolytic viruses including herpes simplex virus, adenovirus, and vaccinia virus—have demonstrated safety and efficacy for treating melanoma, glioma, pancreatic cancer, and other malignancies. Combination of oncolytic virotherapy with other therapeutic strategies, including immune checkpoint inhibitors, has been shown to increase treatment effectiveness and patient survival in select cases.
9.1. Implications for Future Cancer Therapy
Current oncolytic virotherapy development in cancer treatment is poised to revolutionize cancer therapeutics, particularly in precision medicine contexts. It is feasible to advance OVs to specifically target particular tumor types while simultaneously reprogramming immune systems. Oncolytic viruses should be combined with other immunotherapies, including checkpoint inhibitors, as they enable the immune system to detect cancer cells. Biomarkers for evaluating patient status following oncolytic virotherapy and improving viral selectivity represent important areas for development.
9.2. Final Comments on Oncolytic Virotherapy Future
Oncolytic virotherapy represents a novel form of cancer treatment with substantial potential effectiveness. Its capacity to directly induce cancer cell death while modulating tumor conditions and stimulating systemic immune responses constitutes a unique feature. However, questions remain regarding: delivery system optimization, viral adaptability, and patient response variability. Further research and clinical trials are essential to discover additional oncolytic virotherapy potential and establish its role as a cornerstone of oncological cancer treatment. As understanding of relationships between oncolytic viruses and immune systems deepens, the future of oncolytic virotherapy as a platform for superior and selective cancer treatment appears increasingly promising.
Acknowledgements
Funding
This research project is funded in part by Universiti Malaya private funding under the project code IF038-2025. Funding for this research was provided by the National Natural Science Foundation of China [grant No.82201897]; the Science and technology innovation Commission of Shenzhen [grant numbers JCYJ20210324130005013]; the Health and Medical Scientific Research Project of Shenzhen Bao'an Medical Association [grant numbers BAYXH2024027].
Author contributions
QT, BN and KWL conceived and designed the review. SC, CLY and BY drafted the manuscript. SC and BP provided expert clinical consultation and critical suggestions. YQZ prepared the original artwork. PVN contributed to manuscript revision. All authors read and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Russell SJ, Peng KW, Bell JC. Oncolytic virotherapy. Nature Biotechnology. 2012Jul;30(7):658-70
2. Wei D, Xu J, Liu XY, Chen ZN, Bian H. Fighting cancer with viruses: oncolytic virus therapy in China. Human Gene Therapy. 2018Feb1;29(2):151-9
3. Zheng M, Huang J, Tong A, Yang H. Oncolytic viruses for cancer therapy: barriers and recent advances. Molecular Therapy-Oncolytics. 2019Dec20;15:234-47
4. Woller N, Gürlevik E, Ureche CI, Schumacher A, Kühnel F. Oncolytic viruses as anticancer vaccines. Frontiers in oncology. 2014Jul21;4:188
5. Goradel NH, Alizadeh A, Hosseinzadeh S, Taghipour M, Ghesmati Z, Arashkia A, Negahdari B. Oncolytic virotherapy as promising immunotherapy against cancer: mechanisms of resistance to oncolytic viruses. Future Oncology. 2022Jan1;18(2):245-59
6. Vähä-Koskela MJ, Heikkilä JE, Hinkkanen AE. Oncolytic viruses in cancer therapy. Cancer letters. 2007Sep8;254(2):178-216
7. Zeyaullah M, Patro M, Ahmad I, Ibraheem K, Sultan P, Nehal M, Ali A. Oncolytic viruses in the treatment of cancer: a review of current strategies. Pathology & Oncology Research. 2012Oct;18:771-81
8. Cheema TA, Wakimoto H, Fecci PE, Ning J, Kuroda T, Jeyaretna DS, Martuza RL, Rabkin SD. Multifaceted oncolytic virus therapy for glioblastoma in an immunocompetent cancer stem cell model. Proceedings of the National Academy of Sciences. 2013Jul16;110(29):12006-11
9. Kohlhapp FJ, Kaufman HL. Molecular pathways: mechanism of action for talimogene laherparepvec, a new oncolytic virus immunotherapy. Clinical Cancer Research. 2016Mar1;22(5):1048-54
10. Ma J, Ramachandran M, Jin C, Quijano-Rubio C, Martikainen M, Yu D, Essand M. Characterization of virus-mediated immunogenic cancer cell death and the consequences for oncolytic virus-based immunotherapy of cancer. Cell death & disease. 2020Jan22;11(1):48
11. Aurelian L, Bollino D, Colunga A. The oncolytic virus Î PK has multimodal anti-tumor activity. FEMS Pathogens and Disease. 2016Jul1;74(5):ftw050
12. Bartlett DL, Liu Z, Sathaiah M, Ravindranathan R, Guo Z, He Y, Guo ZS. Oncolytic viruses as therapeutic cancer vaccines. Molecular cancer. 2013Dec;12:1-6
13. De Matos AL, Franco LS, McFadden G. Oncolytic viruses and the immune system: the dynamic duo. Molecular therapy Methods & clinical development. 2020Jun12;17:349-58
14. Post DE, Devi NS, Li Z, Brat DJ, Kaur B, Nicholson A, Olson JJ, Zhang Z, Van Meir EG. Cancer therapy with a replicating oncolytic adenovirus targeting the hypoxic microenvironment of tumors. Clinical cancer research. 2004Dec15;10(24):8603-12
15. Law AM, Valdes-Mora F, Gallego-Ortega D. Myeloid-derived suppressor cells as a therapeutic target for cancer. Cells. 2020Feb27;9(3):561
16. Vazaios K, van Berkum RE, Calkoen FG, van der Lugt J, Hulleman E. OV modulators of the paediatric brain TIME: Current status, combination strategies, limitations and future directions. International Journal of Molecular Sciences. 2024May3;25(9):5007
17. Guo ZS, Thorne SH, Bartlett DL. Oncolytic virotherapy: molecular targets in tumor-selective replication and carrier cell-mediated delivery of oncolytic viruses. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer. 2008Apr1;1785(2):217-31
18. Sivanandam V, LaRocca CJ, Chen NG, Fong Y, Warner SG. Oncolytic viruses and immune checkpoint inhibition: the best of both worlds. Molecular Therapy-Oncolytics. 2019Jun28;13:93-106
19. Liang S, Wu YS, Li DY, Tang JX, Liu HF. Autophagy in viral infection and pathogenesis. Frontiers in Cell and Developmental Biology. 2021Oct15;9:766142
20. Inoue H, Tani KJ. Multimodal immunogenic cancer cell death as a consequence of anticancer cytotoxic treatments. Cell Death & Differentiation. 2014Jan;21(1):39-49
21. Twumasi-Boateng K, Pettigrew JL, Kwok YE, Bell JC, Nelson BH. Oncolytic viruses as engineering platforms for combination immunotherapy. Nature Reviews Cancer. 2018Jul;18(7):419-32
22. Liikanen I, Ahtiainen L, Hirvinen ML, Bramante S, Cerullo V, Nokisalmi P, Hemminki O, Diaconu I, Pesonen S, Koski A, Kangasniemi L. Oncolytic adenovirus with temozolomide induces autophagy and antitumor immune responses in cancer patients. Molecular Therapy. 2013Jun1;21(6):1212-23
23. Schäfer S, Weibel S, Donat U, Zhang Q, Aguilar RJ, Chen NG. et al. Vaccinia virus-mediated intra-tumoral expression of matrix metalloproteinase 9 enhances oncolysis of PC-3 xenograft tumors. BMC Cancer. 2012;12:1-9
24. Kleijn A, Kloezeman J, Treffers-Westerlaken E, Fulci G, Leenstra S, Dirven C. et al. The in vivo therapeutic efficacy of the oncolytic adenovirus Delta24-RGD is mediated by tumor-specific immunity. PloS ONE. 2014;9(5):e97495
25. Sette P, Amankulor N, Li A, Marzulli M, Leronni D, Zhang M. et al. GBM-targeted oHSV armed with matrix metalloproteinase 9 enhances anti-tumor activity and animal survival. Molecular Therapy-Oncolytics. 2019;15:214-22
26. Ehrig K, Kilinc MO, Chen NG, Stritzker J, Buckel L, Zhang Q. et al. Growth inhibition of different human colorectal cancer xenografts after a single intravenous injection of oncolytic vaccinia virus GLV-1h68. J Transl Med. 2013;11:1-15
27. Hu Z, Gerseny H, Zhang Z, Chen Y-J, Berg A, Zhang Z, et al.Oncolytic adenovirus expressing soluble TGFβ receptor II-Fc-mediated inhibition of established bone metastases. A safe and effective systemic therapeutic approach for breast cancer. Mol Ther. 2011;19(9):1609-18
28. Kaufman HL, Kim DW, DeRaffele G, Mitcham J, Coffin RS, Kim-Schulze S. Local and distant immunity induced by intralesional vaccination with an oncolytic herpes virus encoding GM-CSF in patients with stage IIIc and IV melanoma. Ann Surg Oncol. 2010;17:718 30
29. Pol JG, Atherton MJ, Stephenson KB, Bridle BW, Workenhe ST, Kazdhan N, McGray AR, Wan Y, Kroemer G, Lichty BD. Enhanced immunotherapeutic profile of oncolytic virus-based cancer vaccination using cyclophosphamide preconditioning. Journal for Immunotherapy of Cancer. 2020Aug13;8(2):e000981
30. Ghouse SM, Nguyen H-M, Bommareddy PK, Guz-Montgomery K, Saha D. Oncolytic herpes simplex virus encoding IL12 controls triple-negative breast cancer growth and metastasis. Front Oncol. 2020;10:384
31. Rajani K, Parrish C, Kottke T, Thompson J, Zaidi S, Ilett L, Shim KG, Diaz RM, Pandha H, Harrington K, Coffey M. Combination therapy with reovirus and anti-PD-1 blockade controls tumor growth through innate and adaptive immune responses. Molecular Therapy. 2016Jan1;24(1):166-74
32. Shalhout SZ, Miller DM, Emerick KS, Kaufman HL. Therapy with oncolytic viruses: progress and challenges. Nature reviews Clinical oncology. 2023Mar;20(3):160-77
33. Wang L, Dunmall LS, Cheng Z, Wang Y. Remodeling the tumor microenvironment by oncolytic viruses: beyond oncolysis of tumor cells for cancer treatment. Journal for immunotherapy of cancer. 2022May31;10(5):e004167
34. Chen CY, Hutzen B, Wedekind MF, Cripe TP. Oncolytic virus and PD-1/PD-L1 blockade combination therapy. Oncolytic virotherapy. 2018Jul;31:65-77
35. Taylor IP, Lopez JA. Oncolytic adenoviruses and the treatment of pancreatic cancer: a review of clinical trials. J Cancer Res Clin Oncol. 2023Aug;149(10):8117-8129
36. Chou J, Kern ER, Whitley RJ, Roizman B. Mapping of herpes simplex virus-1 neurovirulence to gamma 134.5, a gene nonessential for growth in culture. Science. 1990Nov30;250(4985):1262-6
37. Chiocca EA, Rabkin SD. Oncolytic viruses and their application to cancer immunotherapy. Cancer Immunol Res. 2014 Apr;2(4):295-300. Doi: 10.1158/2326-6066.CIR-14-0015. Erratum in: Cancer Immunol Res. 2014Jul;2(7):699
38. Bourhill T, Mori Y, Rancourt DE, Shmulevitz M, Johnston RN. Going (Reo)Viral: Factors Promoting Successful Reoviral Oncolytic Infection. Viruses. 2018Aug11;10(8):421
39. Carew JS, Espitia CM, Zhao W, Kelly KR, Coffey M, Freeman JW, Nawrocki ST. Reolysin is a novel reovirus-based agent that induces endoplasmic reticular stress-mediated apoptosis in pancreatic cancer. Cell Death Dis. 2013Jul18;4(7):e728
40. Chakraborty S. et al. Oncolytic Poxviruses: Immunotherapy and Mechanistic Insights. Journal of Virology. 2018 92(12). Doi: 10.1128/JVI.00756-18
41. Cripe TP, Ngo MC, Geller JI, Louis CU, Currier MA, Racadio JM, Towbin AJ, Rooney CM, Pelusio A, Moon A, Hwang TH, Burke JM, Bell JC, Kirn DH, Breitbach CJ. Phase 1 study of intratumoral Pexa-Vec (JX-594), an oncolytic and immunotherapeutic vaccinia virus, in pediatric cancer patients. Mol Ther. 2015Mar;23(3):602-8
42. Cary ZD, Willingham MC, Lyles DS. Oncolytic vesicular stomatitis virus induces apoptosis in U87 glioblastoma cells by a type II death receptor mechanism and induces cell death and tumor clearance in vivo. J Virol. 2011Jun;85(12):5708-17
43. Puzanov I, Milhem MM, Minor D, Hamid O, Li A, Chen L, Chastain M, Gorski KS, Anderson A, Chou J, Kaufman HL. Talimogene laherparepvec in combination with ipilimumab in previously untreated, unresectable stage IIIB-IV melanoma. Journal of Clinical Oncology. 2016Aug1;34(22):2619-26
44. Cheng PH, Lian S, Zhao R, Rao XM, McMasters KM, Zhou HS. A combination of the autophagy inducer rapamycin and oncolytic adenovirus improves antitumor effect in cancer cells. Virology journal. 2013Dec;10:1-3
45. Ylösmäki E, Hakkarainen T, Hemminki A, Visakorpi T, Andino R, Saksela K. Generation of a conditionally replicating adenovirus based on targeted destruction of E1A mRNA by a cell type-specific microRNA. Journal of Virology. 2008Nov15;82(22):11009-15
46. Heo J, Reid T, Ruo L, Breitbach CJ, Rose S, Bloomston M, Cho M, Lim HY, Chung HC, Kim CW, Burke J. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia JX-594 in liver cancer. Nature medicine. 2013Mar;19(3):329-36
47. Kotterman MA, Schaffer DV. Engineering adeno-associated viruses for clinical gene therapy. Nature Reviews Genetics. 2014Jul;15(7):445-51
48. Engeland CE, Grossardt C, Veinalde R, Bossow S, Lutz D, Kaufmann JK, Shevchenko I, Umansky V, Nettelbeck DM, Weichert W, Jäger D. CTLA-4 and PD-L1 checkpoint blockade enhances oncolytic measles virus therapy. Molecular Therapy. 2014Nov1;22(11):1949-59
49. Guedan S, Rojas JJ, Gros A, Mercade E, Cascallo M, Alemany R. Hyaluronidase expression by an oncolytic adenovirus enhances its intratumoral spread and suppresses tumor growth. Molecular Therapy. 2010Jul1;18(7):1275-83
50. Breitbach CJ, Burke J, Jonker D, Stephenson J, Haas AR, Chow LQ, Nieva J, Hwang TH, Moon A, Patt R, Pelusio A. Intravenous delivery of a multi-mechanistic cancer-targeted oncolytic poxvirus in humans. Nature. 2011Sep1;477(7362):99-102
51. Post DE, Sandberg EM, Kyle MM, Devi NS, Brat DJ, Xu Z, Tighiouart M, Van Meir EG. Targeted cancer gene therapy using a hypoxia-inducible factor-dependent oncolytic adenovirus armed with interleukin-4. Cancer research. 2007Jul15;67(14):6872-81
52. Tian Y, Xie D, Yang L. Engineering strategies to enhance oncolytic viruses in cancer immunotherapy. Signal Transduct Target Ther. 2022Apr6;7(1):117
53. Verheije MH, Rottier PJ. Retargeting of viruses to generate oncolytic agents. Adv Virol. 2012;2012:798526
54. Li Y, Idamakanti N, Arroyo T, Thorne S, Reid T, Nichols S, VanRoey M, Colbern G, Nguyen N, Tam O, Working P, Yu DC. Dual promoter-controlled oncolytic adenovirus CG5757 has strong tumor selectivity and significant antitumor efficacy in preclinical models. Clin Cancer Res. 2005Dec15;11(24 Pt 1):8845-55
55. Lei N, Shen FB, Chang JH, Wang L, Li H, Yang C, Li J, Yu DC. An oncolytic adenovirus expressing granulocyte macrophage colony-stimulating factor shows improved specificity and efficacy for treating human solid tumors. Cancer Gene Ther. 2009Jan;16(1):33-43
56. Muthukutty P, Yoo SY. Oncolytic Virus Engineering and Utilizations: Cancer Immunotherapy Perspective. Viruses. 2023Jul28;15(8):1645
57. Tur-Planells V, García-Sastre A, Cuadrado-Castano S, Nistal-Villan E. Engineering Non-Human RNA Viruses for Cancer Therapy. Vaccines (Basel). 2023Oct20;11(10):1617
58. Zhang W, Ge K, Zhao Q, Zhuang X, Deng Z, Liu L, Li J, Zhang Y, Dong Y, Zhang Y, Zhang S, Liu B. A novel oHSV-1 targeting telomerase reverse transcriptase-positive cancer cells via tumor-specific promoters regulating the expression of ICP4. Oncotarget. 2015Aug21;6(24):20345-55 doi: 10.18632/oncotarget.3884. PMID: 25972362; PMCID: PMC4653009
59. Longo SL, Griffith C, Glass A, Shillitoe EJ, Post DE. Development of an oncolytic herpes simplex virus using a tumor-specific HIF-responsive promoter. Cancer Gene Ther. 2011Feb;18(2):123-34
60. Hou R, Goloubeva O, Neuberg DS, Strominger JL, Wilson SB. Interleukin-12 and interleukin-2 induce invariant natural killer T-cell cytokine secretion and perforin expression independent of T-cell receptor activation. Immunology. 2003Sep;110(1):30-7
61. Ribas A, Dummer R, Puzanov I, VanderWalde A, Andtbacka RH, Michielin O, Olszanski AJ, Malvehy J, Cebon J, Fernandez E, Kirkwood JM. Oncolytic virotherapy promotes intratumoral T cell infiltration and improves anti-PD-1 immunotherapy. Cell. 2017Sep7;170(6):1109-19
62. Chesney J, Puzanov I, Collichio F, Singh P, Milhem MM, Glaspy J, Hamid O, Ross M, Friedlander P, Garbe C, Logan TF. Randomized, open-label phase II study evaluating the efficacy and safety of talimogene laherparepvec in combination with ipilimumab versus ipilimumab alone in patients with advanced, unresectable melanoma. Journal of Clinical Oncology. 2018Jun10;36(17):1658-67
63. Meng S, Su B, Li W, Ding Y, Tang L, Zhou W, Song Y, Li H, Zhou C. Enhanced antitumor effect of novel dual-targeted paclitaxel liposomes. Nanotechnology. 2010Sep17;21(41):415103
64. Yao YD, Sun TM, Huang SY, Dou S, Lin L, Chen JN, Ruan JB, Mao CQ, Yu FY, Zeng MS, Zang JY. Targeted delivery of PLK1-siRNA by ScFv suppresses HER2+ breast cancer growth and metastasis. Science translational medicine. 2012Apr18;4(130):130ra48 -
65. Nosaki K, Hamada K, Takashima Y, Sagara M, Matsumura Y, Miyamoto S, Hijikata Y, Okazaki T, Nakanishi Y, Tani K. A novel, polymer-coated oncolytic measles virus overcomes immune suppression and induces robust antitumor activity. Molecular Therapy-Oncolytics. 2016 Jan 1;3
66. Xu C, Lu Z, Luo Y, Liu Y, Cao Z, Shen S, Li H, Liu J, Chen K, Chen Z, Yang X. Targeting of NLRP3 inflammasome with gene editing for the amelioration of inflammatory diseases. Nature Communications. 2018Oct5;9(1):4092
67. Zamarin D, Palese P. Oncolytic Newcastle disease virus for cancer therapy: old challenges and new directions. Future microbiology. 2012Mar1;7(3):347-67
68. Johnson DB, Puzanov I, Kelley MC. Talimogene laherparepvec (T-VEC) for the treatment of advanced melanoma. Immunotherapy. 2015Jul1;7(6):611-9
69. Nguyen HM, Bommareddy PK, Silk AW, Saha D. Optimal timing of PD-1 blockade in combination with oncolytic virus therapy. InSeminars in cancer biology. 2022 Nov 1 (Vol. 86, pp. 971-980). Academic Press
70. Roberts DM, Nanda A, Havenga MJ, Abbink P, Lynch DM, Ewald BA, Liu J, Thorner AR, Swanson PE, Gorgone DA, Lifton MA. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature. 2006May11;441(7090):239-43
71. Kirn DH, Wang Y, Liang W, Contag CH, Thorne SH. Enhancing poxvirus oncolytic effects through increased spread and immune evasion. Cancer research. 2008Apr1;68(7):2071-5
72. Uchida H, Marzulli M, Nakano K, Goins WF, Chan J, Hong CS, Mazzacurati L, Yoo JY, Haseley A, Nakashima H, Baek H. Effective treatment of an orthotopic xenograft model of human glioblastoma using an EGFR-retargeted oncolytic herpes simplex virus. Molecular Therapy. 2013Mar1;21(3):561-9
73. Zamarin D, Holmgaard RB, Subudhi SK, Park JS, Mansour M, Palese P, Merghoub T, Wolchok JD, Allison JP. Localized oncolytic virotherapy overcomes systemic tumor resistance to immune checkpoint blockade immunotherapy-Science Translational Medicine. 2014 Mar 5;6(226):226ra32-.
74. Zamarin D. et al. Pre-existing immunity to oncolytic viruses: implications for virotherapy. Cancer Immunol Res. 2020;8(11):1305-1312
75. DePeaux K, Delgoffe GM. Integrating innate and adaptive immunity in oncolytic virus therapy. Trends in cancer. 2024Feb1;10(2):135-46
76. Shin DH, Nguyen T, Ozpolat B, Lang F, Alonso M, Gomez-Manzano C, Fueyo J. Current strategies to circumvent the antiviral immunity to optimize cancer virotherapy. Journal for immunotherapy of cancer. 2021Apr1;9(4):e002086
77. Fukuhara H, Ino Y, Todo T. Oncolytic virus therapy: A new era of cancer treatment at dawn. Cancer Sci. 2016Oct;107(10):1373-1379 doi: 10.1111/cas.13027. Epub 2016 Sep 9. PMID: 27486853; PMCID: PMC5084676
78. Ahi YS, Bangari DS, Mittal SK. Adenoviral vector immunity: its implications and circumvention strategies. Curr Gene Ther. 2011Aug;11(4):307-20
79. Wang JH, Gessler DJ, Zhan W, Gallagher TL, Gao G. Adeno-associated virus as a delivery vector for gene therapy of human diseases. Signal Transduct Target Ther. 2024Apr3;9(1):78
80. Agelidis AM, Shukla D. Cell entry mechanisms of HSV: what we have learned in recent years. Future Virol. 2015Oct1;10(10):1145-1154
81. Carfí A, Willis SH, Whitbeck JC, Krummenacher C, Cohen GH, Eisenberg RJ, Wiley DC. Herpes simplex virus glycoprotein D bound to the human receptor HveA. Mol Cell. 2001Jul;8(1):169-79
82. Todo T, Rabkin SD, Chahlavi A, Martuza RL. Corticosteroid administration does not affect viral oncolytic activity but inhibits antitumor immunity in replication-competent herpes simplex virus tumor therapy. Hum Gene Ther. 1999Nov20;10(17):2869-78
83. Wan PK, Fernandes RA, Seymour LW. Oncolytic viruses and antibodies: are they more successful when delivered separately or when engineered as a single agent? J Immunother Cancer. 2023Aug;11(8):e006518
84. Ju F, Luo Y, Lin C, Jia X, Xu Z, Tian R, Lin Y, Zhao M, Chang Y, Huang X, Li S, Ren W, Qin Y, Yu M, Jia J, Han J, Luo W, Zhang J, Fu G, Ye X, Huang C, Xia N. Oncolytic virus expressing PD-1 inhibitors activates a collaborative intratumoral immune response to control tumor and synergizes with CTLA-4 or TIM-3 blockade. J Immunother Cancer. 2022 Jun;10(6): e004762. doi: 10.1136/jitc-2022-004762. Erratum in: J Immunother Cancer. 2024Jan30;12(1):e004762corr1
85. Elkashif A, Alhashimi M, Sayedahmed EE, Sambhara S, Mittal SK. Adenoviral vector-based platforms for developing effective vaccines to combat respiratory viral infections. Clin Transl Immunology. 2021Oct12;10(10):e1345
86. Nidetz NF, Gallagher TM, Wiethoff CM. Inhibition of type I interferon responses by adenovirus serotype-dependent Gas6 binding. Virology. 2018Feb;515:150-157
87. Ma W, He H, Wang H. Oncolytic herpes simplex virus and immunotherapy. BMC Immunol. 2018Dec18;19(1):40 doi: 10.1186/s12865-018-0281-9. PMID: 30563466; PMCID: PMC6299639
88. Naumenko V, Rajwani J, Turk M, Zhang C, Tse M, Davis RP, Kim D, Rakic A, Dastidar H, Van S, Mah LK, Kaul EK, Chekhonin VP, Mahoney DJ, Jenne CN. Repeated dosing improves oncolytic rhabdovirus therapy in mice via interactions with intravascular monocytes. Commun Biol. 2022Dec19;5(1):1385
89. Melcher A. Oncolytic Virotherapy: Single Cycle Cures or Repeat Treatments? (Repeat Dosing Is Crucial!). Mol Ther. 2018Aug1;26(8):1875-1876
90. Wu B, Zhang B, Li B, Wu H, Jiang M. Cold and hot tumors: from molecular mechanisms to targeted therapy. Signal Transduct Target Ther. 2024Oct18;9(1):274
91. Ponterio E, Haas TL, De Maria R. Oncolytic virus and CAR-T cell therapy in solid tumors. Front Immunol. 2024Oct30;15:1455163 doi: 10.3389/fimmu.2024.1455163. PMID: 39539554; PMCID: PMC11557337
92. Jin KT, Du WL, Liu YY, Lan HR, Si JX, Mou XZ. Oncolytic Virotherapy in Solid Tumors: The Challenges and Achievements. Cancers (Basel). 2021Feb3;13(4):588
93. Belcaid Z, Berrevoets C, Choi J, van Beelen E, Stavrakaki E, Pierson T, Kloezeman J, Routkevitch D, van der Kaaij M, van der Ploeg A, Mathios D, Sleijfer S, Dirven C, Lim M, Debets R, Lamfers MLM. Low-dose oncolytic adenovirus therapy overcomes tumor-induced immune suppression and sensitizes intracranial gliomas to anti-PD-1 therapy. Neurooncol Adv. 2020Feb3;2(1):vdaa011
94. Bartee E. Potential of oncolytic viruses in the treatment of multiple myeloma. Oncolytic Virother. 2018Feb23;7:1-12
95. Thirukkumaran CM, Morris DG. Oncolytic virotherapy for multiple myeloma: past, present, and future. Bone Marrow Res. 2011;2011:632948
96. Yang C, Hua N, Xie S, Wu Y, Zhu L, Wang S, Tong X. Oncolytic viruses as a promising therapeutic strategy for hematological malignancies. Biomed Pharmacother. 2021Jul;139:111573
97. Rehman H, Silk AW, Kane MP, Kaufman HL. Into the clinic: Talimogene laherparepvec (T-VEC), a first-in-class intratumoral oncolytic viral therapy. J Immunother Cancer. 2016Sep20;4:53
98. Bommareddy PK, Patel A, Hossain S, Kaufman HL. Talimogene Laherparepvec (T-VEC) and Other Oncolytic Viruses for the Treatment of Melanoma. Am J Clin Dermatol. 2017Feb;18(1):1-15
99. Doniņa S, Strēle I, Proboka G, Auziņš J, Alberts P, Jonsson B, Venskus D, Muceniece A. Adapted ECHO-7 virus Rigvir immunotherapy (oncolytic virotherapy) prolongs survival in melanoma patients after surgical excision of the tumour in a retrospective study. Melanoma Res. 2015Oct;25(5):421-6
100. Matsuzaka Y, Yashiro R. Advances in Gene Therapy with Oncolytic Viruses and CAR-T Cells and Therapy-Related Groups. Curr Issues Mol Biol. 2025Apr10;47(4):268 doi: 10.3390/cimb47040268. PMCID: PMC12025996
101. Zou H, Mou XZ, Zhu B. Combining of Oncolytic Virotherapy and Other Immunotherapeutic Approaches in Cancer: A Powerful Functionalization Tactic. Glob Chall. 2022Oct20;7(1):2200094
102. Li Q, Tan F, Wang Y, Liu X, Kong X, Meng J, Yang L, Cen S. The gamble between oncolytic virus therapy and IFN. Front Immunol. 2022Aug25;13:971674
103. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021Feb;20(2):101-124
104. Zheng M, Huang J, Tong A, Yang H. Oncolytic Viruses for Cancer Therapy: Barriers and Recent Advances. Mol Ther Oncolytics. 2019Nov2;15:234-247
105. Ageenko A, Vasileva N, Richter V, Kuligina E. Combination of Oncolytic Virotherapy with Different Antitumor Approaches against Glioblastoma. Int J Mol Sci. 2024Feb7;25(4):2042
106. Zhang T, Jou TH, Hsin J, Wang Z, Huang K, Ye J, Yin H, Xing Y. Talimogene laherparepvec (T-VEC): a review of the recent advances in cancer therapy. Journal of clinical medicine. 2023Jan31;12(3):1098
107. Lovatt C, Parker AL. Oncolytic Viruses and Immune Checkpoint Inhibitors: The "Hot" New Power Couple. Cancers (Basel). 2023Aug19;15(16):4178 Doi: 10.3390/cancers15164178. PMID: 37627206; PMCID: PMC10453115
108. Guo ZS, Bartlett DL. Oncolytic viruses as a platform for multimodal cancer therapeutics: a promising land. Cancer Gene Ther. 2014Jul;21(7):261-3
109. Wang Z, Sun P, Li Z, Xiao S. Clinical Advances and Future Directions of Oncolytic Virotherapy for Head and Neck Cancer. Cancers (Basel). 2023Nov4;15(21):5291
110. Yan Z, Zhang Z, Chen Y, Xu J, Wang J, Wang Z. Enhancing cancer therapy: the integration of oncolytic virus therapy with diverse treatments. Cancer Cell Int. 2024Jul11;24(1):242 doi: 10.1186/s12935-024-03424-z. PMID: 38992667; PMCID: PMC11238399
Author contact
![]() Corresponding authors: Prof. Quan Tang, Research Laboratory, Shenzhen Baoan Women's and Children's Hospital, Shenzhen, Guangdong, China, ORCID: 0009-0002-3870-8082, Mail: tangquan1987com. Prof. Bommireddy Naveen, Department of Chemical and Biological Engineering, Gachon University, Seongnam-Si, Gyeonggi-do, Republic of Korea, Mail: naveenbac.kr. Prof. Khin Wee Lai, Department of Biomedical Engineering, Faculty of Engineering, Universiti Malaya, Kuala Lumpur, 50603, Malaysia, Email: lai.khinweeedu.my.
Corresponding authors: Prof. Quan Tang, Research Laboratory, Shenzhen Baoan Women's and Children's Hospital, Shenzhen, Guangdong, China, ORCID: 0009-0002-3870-8082, Mail: tangquan1987com. Prof. Bommireddy Naveen, Department of Chemical and Biological Engineering, Gachon University, Seongnam-Si, Gyeonggi-do, Republic of Korea, Mail: naveenbac.kr. Prof. Khin Wee Lai, Department of Biomedical Engineering, Faculty of Engineering, Universiti Malaya, Kuala Lumpur, 50603, Malaysia, Email: lai.khinweeedu.my.