Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2026; 17(7):1265-1273. doi:10.7150/jca.134391 This issue Cite
Research Paper
Association of DNA damage response gene polymorphisms with treatment response and prognosis in acute myeloid leukemia
Amin Zhang1,2#, Hanyang Wu1#, Wancheng Liu3 ![]() , Daoxin Ma1,4,5
, Daoxin Ma1,4,5 ![]()
1. Department of Hematology, Qilu Hospital of Shandong University, No.107, West of Wenhua Road, Jinan, 250012, Shandong, People's Republic of China.
2. Department of Pediatrics, Qilu Hospital of Shandong University, No.107, West of Wenhua Road, Jinan, 250012, Shandong, People's Republic of China.
3. Department of Clinical Laboratory, Qilu Hospital of Shandong University, No.107, West of Wenhua Road, Jinan, 250012, Shandong, People's Republic of China.
4. Shandong Key Laboratory of Hematological Diseases and Immune Microenvironment, Qilu Hospital of Shandong University, No.107, West of Wenhua Road, Jinan, 250012, Shandong, People's Republic of China.
5. Shandong Provincial Clinical Research Center for Hematological Diseases, Qilu Hospital of Shandong University, No.107, West of Wenhua Road, Jinan, 250012, Shandong, People's Republic of China.
# These authors contributed equally to this work.
Received 2026-3-13; Accepted 2026-5-13; Published 2026-7-1
Abstract

The potential functional Single Nucleotide Polymorphism (SNPs) of DNA Damage Response (DDR)-related genes may influence AML aggressiveness and clinical outcomes through regulating genomic stability. However, systematic evaluation of functionally relevant SNPs across the entire DDR cascade in AML remains limited. In our study, significant associations were identified across multiple DDR modules. In the damage sensing module, MRE11 rs2155209 TC/CC genotypes were protective against hyperleukocytosis and associated with longer OS, while NBS1 rs1805794 CG/GG genotypes increased hyperleukocytosis risk. In the signal transduction module, CHEK1 rs521102 AA genotype was a risk factor for thrombocytopenia but unexpectedly associated with prolonged OS; CHEK2 rs738722 TT genotype increased hyperleukocytosis risk; RAD17 rs1045051 GT/GG genotype conferred a higher relapse risk; and BRCA1 rs1799949 GA/AA genotype as a protective factor for high BM blast percentage. In the DNA repair module, LIG3 rs3744356 TT genotypes predicted higher WBC counts and lower complete remission rate. Both LIG3 rs3744356 TT genotype and MGMT rs2296675 GA genotype were associated with shorter OS (HR = 1.719, 95% CI = 1.082 -2.741, p = 0.022; HR = 1.511, 95% CI = 1.046 - 2.182, p = 0.028). SNPs in effector genes TP53 and CDKN1A showed no significant associations. In summary, SNPs across the DDR pathway—particularly in damage sensing, signal transduction, and DNA repair modules—are significantly associated with AML clinical features, treatment response, and survival. Our findings provide a foundation for developing DDR-based polygenic risk models and support the potential integration of germline SNP profiling into precision medicine strategies for AML.
Keywords: AML, SNPs, DDR pathway, treatment response
Introduction
Acute myeloid leukemia (AML) is a common hematological malignancy driven by the abnormal accumulation of genomic mutations in hematopoietic stem/progenitor cells [1, 2]. Although the combination of cytarabine and daunorubicin has been established as the conventional standard-intensity chemotherapy regimen for AML, approximately 20 - 40% of patients fail to achieve complete remission (CR) [3]. The main mechanism of conventional chemotherapy is to induce DNA damage of AML cells, thereby triggering the cell apoptosis. The levels of DNA damage could influence the effects of chemotherapy [4, 5]. Therefore, it is of importance to explore the intrinsic DNA damage of AML cells to evaluate the effects of chemotherapy.
Genetic variations have been reported to affect the chemosensitivity or DNA damage of AML cells at different aspects [6]. For the somatic variations, acquired driver mutations directly lead to differentiation arrest and proliferative advantage acquisition in AML cells [7-9]. Therefore, somatic mutations in CEBPA, NPM1 and FLT3 et al. have been incorporated into the risk stratification of AML by the European LeukemiaNet (ELN) guidelines, directly guiding prognostic assessment and treatment decisions [10]. For the germline variations, single nucleotide polymorphisms (SNPs) constitute the genetic background underlying interindividual differences, subtly shaping physiological state by influencing mechanisms such as gene transcription, mRNA stability, or protein function. In particular, SNPs in key genes of the DNA damage response (DDR) pathway have been shown to play important roles in prostate cancer, breast cancer to process genotoxic drugs, thereby affecting the sensitivity to chemotherapy [11, 12]. However, given that individual DDR SNPs have been sporadically studied in AML, a systematic, pathway-based evaluation covering the entire signaling cascade, from damage sensing to DNA repair execution, remains lacking.
In this study, we focus on a set of potentially functional SNPs within the DNA damage checkpoint pathway. These loci comprehensively cover the entire signaling cascade, ranging from damage sensing (e.g., sites related to MRE11, RAD50, and NBS1, components of the MRN complex), through signal transduction and amplification (e.g., CHEK2, CHEK1, RAD17), to the central decision point (the TP53 codon 72 polymorphism). Additionally, we include relevant loci for LIG3, which affects end-joining repair, and for the growth factor IGF1, which may interfere with the p53 pathway. This investigation aims to elucidate how this multi-gene SNP profile collectively shapes the clinical presentation of AML and modulates patient response and prognosis following DNA damage-based chemotherapy.
Materials and Methods
Study Population
A total of 245 patients with newly diagnosed AML were enrolled at Qilu Hospital of Shandong University between January 2010 and January 2023. Patients were excluded if they were: 1) under 14 years of age; 2) had received prior anti-leukemic therapy; 3) presented with other primary malignancies; 4) lacked available biological samples. All diagnoses were confirmed according to the National Comprehensive Cancer Network (NCCN) guidelines and the World Health Organization (WHO) classification. The cohort had a median age of 52 years (range: 15 - 79). All patients received induction chemotherapy consisting of an anthracycline combined with cytarabine. Written informed consent was obtained from each participant prior to inclusion in accordance with the Declaration of Helsinki, and the study was approved by the Institutional Ethics Committee of Qilu Hospital (Approval No. KYLL-202204-059). Basic information and clinical characteristics of the patients are summarized in Table 1.
The selected genes and SNPs.
| Genes | SNPs | MAF | HWE (p-value) | |
|---|---|---|---|---|
| DNA damage sensing | MRE11 | rs2155209 | 0.237 | 0.602 |
| NBS1 | rs1805794 | 0.425 | 0.663 | |
| RAD50 | rs2706347 | 0.155 | 0.598 | |
| Signal transduction | ATM | rs1801516 | 0.039 | 0.523 |
| ATR | rs1802904 | 0.040 | 0.336 | |
| CHEK1 | rs521102 | 0.384 | 0.077 | |
| CHEK2 | rs738722 | 0.279 | 0.757 | |
| RAD17 | rs1045051 | 0.368 | 0.744 | |
| BRCA1 | rs1799949 | 0.40 | 0.749 | |
| BARD1 | rs6435862 | 0.102 | 0.754 | |
| Hus1 | rs1056663 | 0.443 | 0.003 | |
| Effector activation | TP53 | rs1042522 | 0.435 | 0.795 |
| P21 | rs1801270 | 0.447 | 0.0009 | |
| DNA repair / Damage resolution | LIG4 | rs1805388 | 0.194 | 0.72 |
| LIG3 | rs3744356 | 0.323 | 0.602 | |
| LIG1 | rs20579 | 0.115 | 0.887 | |
| MGMT | rs2296675 | 0.105 | 0.067 | |
| BRCA2 | rs9567552 | 0.312 | 0.068 |
DNA extraction and SNP genotyping
Peripheral blood samples collected at the time of diagnosis were used for genomic DNA extraction. Mononuclear cells were first isolated by Ficoll-Hypaque density gradient centrifugation, and genomic DNA was subsequently extracted using a commercial kit (TianGen, Beijing, China) according to the manufacturer's protocol. DNA concentration and purity were assessed by spectrophotometry, after which aliquots were stored at -80°C until further processing. Candidate SNPs involved in DDR were curated for this study (Table 2). Selection and functional annotation of these SNPs were based on the dbSNP and UCSC databases. For each SNP, corresponding primers were developed using Agena Assay Designer 4.0. Three primers, one universal extension primer and two amplification primers, were synthesized per locus and purified via polyacrylamide gel electrophoresis (PAGE). Genotyping assays were conducted with approximately 20 ng of input DNA per sample. Detection was performed on a MassARRAY platform, followed by sample analysis using MALDI-TOF/TOF mass spectrometry. To ensure assay accuracy, 6 negative controls (no DNA template) and 6 positive controls (samples with known genotypes) were inter-spiked among the study samples. Additionally, 16 randomly selected samples were genotyped in duplicate across two independent assay panels, yielding a reproducibility rate of 99%.
Clinical features of patients with AML.
| Variable | Case n (%) |
|---|---|
| Gender | |
| Male | 127 (51.8) |
| Female | 118 (48.2) |
| Age (years, median, range) | |
| Median | 52 (15 - 79) |
| < 60 | 172 (70.2) |
| ≥ 60 | 73 (29.8) |
| WBC (×109/L) | |
| Median | 14.74 (0.48 - 351.95) |
| < 100 | 220 (89.8) |
| ≥ 100 | 25 (10.2) |
| PLT (×109/L) | |
| Median | 43 (5 - 861) |
| ≥ 50 | 104 (42.5) |
| < 50 | 141 (57.5) |
| HGB (g/L) | |
| Median | 75 (27 - 136) |
| ≥ 60 | 202 (82.4) |
| < 60 | 43 (17.6) |
| Risk stratification | |
| Favorable | 68 (27.8) |
| Intermediate | 147 (60.0) |
| Adverse | 30 (12.2) |
| Response after 1 cycle of cytarabine- and anthracycline- containing induction therapy | |
| CR | 151 (61.6) |
| No CR | 94 (38.4) |
Statistical analysis
Hardy-Weinberg equilibrium (HWE) was assessed using the chi-square test. SNPs conforming to HWE (p > 0.05) and exhibiting a minor allele frequency (MAF) greater than 5% in the general population were retained for subsequent analysis. Genotyping data were evaluated under four genetic models: dominant, recessive, codominant, and allelic. To assess associations between SNP genotypes/alleles and clinical outcomes, we performed statistical analyses in two stages. First, categorical variables were screened using the chi-square test or Fisher's exact test, as appropriate. Second, univariate binary logistic regression models—adjusted for sex and age—were applied to calculate odds ratios (ORs) with corresponding 95% confidence intervals (95% CIs) and p-values. Given the exploratory nature of multiple SNP association testing across several genetic models and clinical endpoints, we applied the Benjamini-Hochberg procedure to control the false discovery rate (FDR). Adjusted p-values (q-values) were calculated for primary SNP-outcome associations. Findings with nominal p < 0.05 were reported, but emphasis was placed on associations that remained significant after FDR correction.
Survival analysis was performed using Kaplan-Meier curves with comparisons based on the log-rank test. Associations between SNPs and overall survival (OS) in AML patients were evaluated with the Cox proportional hazards model, yielding hazard ratios (HRs) and corresponding 95% CIs. Prognostic variables were initially screened via univariate Cox analysis, and significant factors were subsequently incorporated into a multivariate Cox model for further assessment.
A multivariate Cox regression model with backward selection was applied to assess the independent prognostic significance of the following covariates: age (< 60 vs. ≥ 60 years), ELN risk category (favorable, intermediate, adverse), white blood cell count at diagnosis (< 100 vs. ≥ 100 × 10⁹/L) and PLT count (< 50 vs. ≥ 50 × 10⁹/L) All statistical analyses were conducted using SPSS version 26.0 (IBM Corp., Chicago, IL, USA). A two-sided p-value < 0.05 was considered statistically significant.
Results
SNPs selection and study populations
The selected SNPs of genes associated with DR checkpoint are listed in Table 1. HWE and MAF were used for the initial screening of candidate SNPs. SNPs with p ≥ 0.05 in the HWE test and MAF ≥ 0.05 in the general population were selected for further analysis. Accordingly, two loci with a minor allele frequency (MAF) < 0.05 (ATM rs1801516 and ATR rs1802904) and two loci deviating from the Hardy-Weinberg equilibrium (HWE) (HUS1 rs1056663 and P21 rs1801270) were excluded from further analysis. A total of 245 patients were recruited in this study. The basic information, clinical characteristics and initial treatment response using cytarabine and doxorubicin of these patients were shown in Table 2.
Association between bone marrow (BM) blasts at initial diagnosis
To further explore the value of these SNPs in AML, we first analyzed the relationship between SNPs and the percentage of BM blasts at initial diagnosis (Table 3). The proportion of BM blasts at initial diagnosis served as a surrogate for disease burden. A BM blast percentage of 70% or greater was defined as high, and below 70% as low. The LIG3 rs3744356 and BRCA1 rs1799949 under dominant models, were significantly associated with the percentage of BM blast at initial diagnosis (p < 0.05). After the adjustment of age and sex, the CT genotype under co-dominant model (OR = 2.114, 95% CI = 1.206-3.705, p = 0.009) and CT/TT genotype under dominant model (OR = 1.831, 95% CI = 1.087 - 3.082, p = 0.023) of LIG3 rs3744356 tended to be a risk factor. In contrast, for BRCA1 rs1799949, the GA genotype under the co-dominant model (OR = 0.556, 95% CI = 0.311 - 0.992, p = 0.048) and the GA/AA genotype under the dominant model (OR = 0.539, 95% CI = 0.312 - 0.933, p = 0.027) emerged as protective factors against a higher BM blast percentage at diagnosis.
Association between SNPs and the percentage of BM blast of AML patients.
| Gene | SNP | Model | Genotype | BM blast < 70% | BM blast ≥ 70% | χ2 test p-value | OR (95% CI) | Adjusted p-value |
|---|---|---|---|---|---|---|---|---|
| LIG3 | rs3744356 | Co-dominant | CC | 57 | 55 | 0.02 | ||
| CT | 33 | 69 | 2.114(1.206-3.705) | 0.009 | ||||
| TT | 13 | 14 | 1.087(0.465-2.54) | 0.847 | ||||
| Dominant | CC | 57 | 55 | 0.019 | ||||
| CT/TT | 46 | 83 | 1.831(1.087-3.082) | 0.023 | ||||
| BRCA1 | rs1799949 | Co-dominant | GG | 29 | 58 | 0.065 | ||
| GA | 58 | 62 | 0.556(0.311-0.992) | 0.048 | ||||
| AA | 19 | 19 | 0.723(0.486-1.076) | 0.11 | ||||
| Dominant | GG | 29 | 58 | 0.02 | ||||
| GA/AA | 77 | 81 | 0.539(0.312-0.933) | 0.027 |
Association between peripheral blood characteristics and SNPs
Considering that peripheral blood characteristics might be related to SNPs, we analyzed the association of selected SNPs with WBC, HGB, and PLT in AML patients using the χ2 test or Fisher's exact test. We found the high WBC group consisted of patients with a WBC count equal to or greater than 100 × 109/L, while the low WBC group included patients with a WBC count less than 100 × 109/L. The high PLT group was defined as patients with PLT levels greater or equal to than 50 × 109/L, while the low PLT group included patients with PLT levels less than 50 × 109/L. The high HGB group included patients with an HGB level equal to or greater than 60 g/L, while the low HGB group included patients with an HGB level less than 60 g/L. As shown in Table 3, MRE11 rs2155209 under the co-dominant and dominant models, CHEK2 rs738722 under the co-dominant and recessive models, LIG3 rs3744356 under the recessive model, and NBS1 rs1805794 under the dominant model were significantly associated with AML patients with a high WBC count (p < 0.05). After adjusting for age and sex, the TC genotype of MRE11 rs2155209 under the co-dominant model (OR = 0.29, 95% CI = 0.094 - 0.895, p = 0.031) and the TC/CC genotype under the dominant model (OR = 0.321, 95% CI = 0.114 - 0.899, p = 0.031) were identified as protective factors against high WBC count in AML patients. In contrast, the TT genotype under the codominant model and recessive model of CHEK2 rs738722 are associated with an increased risk of high WBC count in AML. TT genotype in LIG3 rs3744356 recessive model and CG/GG genotype in NBS1 rs1805794 dominant model tended to be a risk factor for AML patients with a high WBC blasts at diagnosis (OR > 1, p < 0.05). However, no SNPs were found to be associated with HGB levels (p > 0.05). After adjustment of sex and age, the AA genotype of CHEK1 in rs521102 is a risk of low PLT under recessive model (OR = 2.188, 95% CI = 1.03 - 4.648, p = 0.042) (Table 4).
The relationship between peripheral blood characteristics and SNPs in AML patients at the first diagnosis.
| Gene | SNP | Model | Genotype | WBC <100×109/L | WBC ≥100×109/L | χ2 testp-value | OR (95% CI) | Adjusted p-value |
| MRE11 | rs2155209 | Co-dominant | TT | 122 | 19 | |||
| TC | 80 | 4 | 0.098 | 0.29 (0.094-0.895) | 0.031 | |||
| CC | 14 | 1 | 0.659 (0.231-1.879) | 0.436 | ||||
| Dominant | TT | 122 | 19 | |||||
| TC/CC | 94 | 5 | 0.032 | 0.321 (0.114-0.899) | 0.031 | |||
| LIG3 | rs3744356 | Recessive | CC/CT | 195 | 19 | |||
| TT | 21 | 6 | 0.032 | 3.019 (1.078-8.454) | 0.035 | |||
| CHEK2 | rs738722 | Co-dominant | CC | 118 | 9 | |||
| CT | 85 | 11 | 0.044 | 1.766 (0.697-4.478) | 0.231 | |||
| TT | 15 | 5 | 2.049 (1.104-3.803) | 0.023 | ||||
| Recessive | CC/CT | 203 | 20 | |||||
| TT | 15 | 5 | 0.041 | 3.336 (1.086-10.25) | 0.035 | |||
| NBS1 | rs1805794 | Dominant | CC | 77 | 4 | |||
| CG/GG | 138 | 21 | 0.047 | 3.099 (1.019-9.421) | 0.046 | |||
| Gene | SNP | Model | Genotype | PLT ≥ 50×109/L | PLT < 50×109/L | χ2 test p-value | OR (95% CI) | Adjusted p-value |
| CHEK1 | rs521102 | Recessive | GG/GA | 88 | 105 | |||
| AA | 11 | 30 | 0.027 | 2.188 (1.03-4.648) | 0.042 |
Polymorphisms are related to sensitivity to cytarabine/anthracycline-induced therapy in AML
We analyzed the associations between SNPs and sensitivity to cytarabine- and anthracycline-containing agents. Among the 245 non-M3 AML patients analyzed and underwent bone marrow cytomorphological assessment after the first course of induction with cytarabine- and anthracycline- containing regimens. Preliminary screening with the χ2 test or Fisher's exact test showed that rs3744356 in LIG3 under the co-dominant and recessive model were significantly correlated with response to chemotherapy (p < 0.05). After adjustment of sex and age, and applying FDR correction, the TT genotype of rs3744356 were found to be significantly related to response to chemotherapy under the co-dominant (OR = 1.81, 95% CI = 1.16 - 2.28, p = 0.009) and recessive models (OR = 3.174, 95% CI = 1.376 - 7.323, p = 0.007), with a decrease in the CR group (Table 5).
Association between SNP of and AML induction therapy response.
| Gene | SNP | Model | Genotype | CR | No CR | χ2 test p-value | OR (95% CI) | Adjusted p-value |
|---|---|---|---|---|---|---|---|---|
| LIG3 | rs3744356 | Co-dominant | CC | 72 | 40 | |||
| CT | 67 | 35 | 0.019 | 0.937 (0.531-1.653) | 0.822 | |||
| TT | 10 | 17 | 1.81 (1.16-2.28) | 0.009 | ||||
| Recessive | CC/CT | 139 | 75 | 0.005 | ||||
| TT | 10 | 17 | 3.174 (1.376-7.323) | 0.007 |
DDR-related polymorphisms are related to AML relapse
Preliminary screening using the χ2 or Fisher's exact test was conducted under three genetic models. As shown in Table 6, RAD17 rs1045051 under co-dominant and dominant models were significantly associated with the AML relapse (p < 0.05). After adjustment of sex and age and applying FDR correction, the GT genotype of RAD17 rs1045051 under co-dominant model (OR = 2.92, 95% CI = 1.347 - 6.327 p = 0.007) and GT/GG genotype under dominant model (OR = 2.433 95% CI = 1.216 - 4.869, p = 0.012) tended to be a risk factor for AML relapse.
Association between SNPs and AML relapse.
| Gene | SNP | Model | Genotype | Relapse | No relapse | χ2 test p-value | OR (95% CI) | Adjusted p-value |
|---|---|---|---|---|---|---|---|---|
| RAD17 | rs1045051 | Co-dominant | TT | 56 | 26 | |||
| GT | 74 | 13 | 0.035 | 2.92 (1.347-6.327) | 0.007 | |||
| GG | 20 | 7 | 1.177 (0.703-1.971) | 0.535 | ||||
| Dominant | TT | 56 | 26 | |||||
| GT/GG | 94 | 20 | 0.021 | 2.433 (1.216-4.869) | 0.012 |
DDR-related polymorphisms are associated with AML overall survival
We then analyzed the relationships between various SNPs and overall survival (OS) in AML patients. Kaplan-Meier analysis revealed an association between the genotype frequency of rs2155209 in MRE11 and prognosis under a dominant model (p < 0.05). Specifically, AML patients carrying the TC/CC genotype exhibited significantly longer overall survival (OS) compared to those with the TT genotype. Kaplan-Meier screening revealed that rs3744356 in LIG3 and rs521102 in CHEK1 under recessive model and rs2296675 in MGMT under Co-dominant model were associated with prognosis (p < 0.05, Figure 1).
Overall survival of AML patient with SNPs of 4 DDR related genes. (A) Rs2155209 SNP and OS in dominant model; (B) Rs3744356 and OS in recessive model; (C) Rs2296675 and OS in co-dominant model; (D) Rs521102 and OS in recessive model.
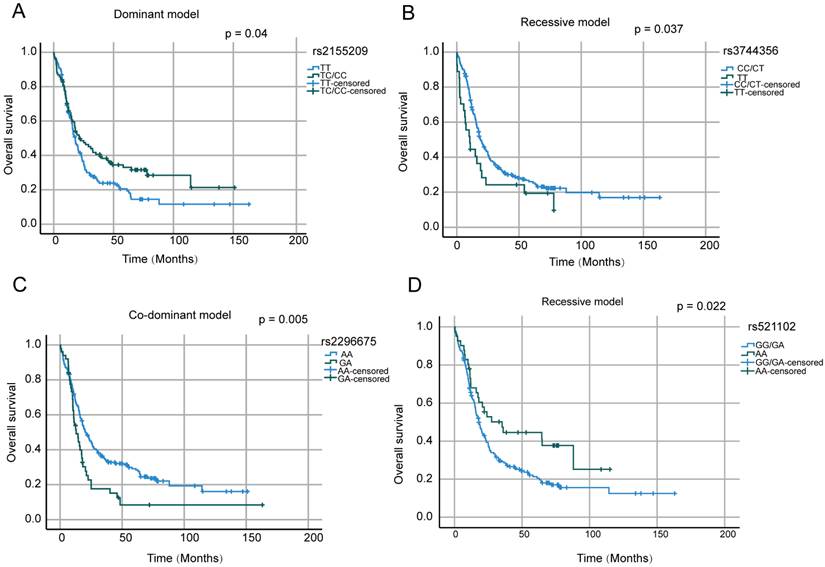
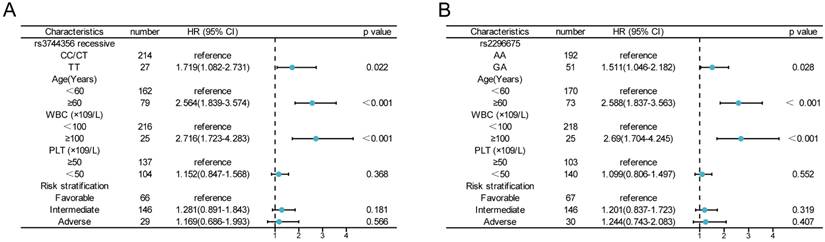
A Cox proportional hazards model with multivariate analysis for OS was used to analyze the above four SNPs, age, risk stratification, WBC and PLT count at diagnosis (Figure 2). In our study, age ≥ 60 years, WBC ≥100×109/L had an independent negative impact on OS. Patients with the TT genotype of rs3744356 in LIG3, remained significantly associated with worse OS (HR = 1.719, 95% CI = 1.082 -2.741, p = 0.022). Similarly, GA genotype of rs2296675 in MGMT remained significantly associated with worse OS (HR = 1.511, 95% CI = 1.046 - 2.182, p = 0.028). These results demonstrated that the TT genotype of rs3744356 in LIG3 and GA genotype of rs2296675 in MGMT is an independent risk prognostic factor for AML patients.
The impact of LIG3 rs3744356 and MGMT rs2296675 on the outcomes of AML patients. (A) Multivariate cox regression analysis of rs3744356 recessive model and clinical characteristics associated with OS; (B) Multivariate cox regression analysis of MGMT rs2296675 co-dominant model and clinical characteristics associated with OS.
Discussion
The development and progression of AML rely on acquisition of specific somatic mutations [13]. SNPs in genes involved in the DDR pathway modulate genomic stability, thereby creating a favorable genetic background for the emergence of driver mutations and subsequent clonal evolution [13-15]. In this study, we systematically evaluated potentially functional SNPs across 4 key modules of the DDR pathway—damage sensing, signal transduction, effector activation, and DNA repair—in patients with AML. Significant associations were identified between several SNPs and AML aggressiveness or clinical outcome, including variants in DNA damage sensing genes (MRE11 rs2155209, NBS1 rs1805794), signal transduction genes (CHEK1 rs521102, CHEK2 rs738722, RAD17 rs1045051 and BRCA1 rs1799949), and DNA repair genes (LIG3 rs3744356 and MGMT rs2296675). In contrast, SNPs of DDR related genes in effector activation, such as TP53 and CDKN1A, showed no significant association with AML. These findings suggest that the impact of germline genetic variation within the DDR pathway on AML may predominantly exert its influence through the integrated processes of damage recognition, signal amplification, and repair execution.
Why do gene SNPs within different modules of the DDR cascade exert distinct functional and clinical effects? The differential impact of SNPs across DDR modules likely reflects the hierarchical and context-dependent architecture of the DDR cascade. Variants in damage sensing genes may modulate the activation threshold of genomic surveillance, whereas SNPs in signal transduction genes influence checkpoint intensity and cell fate decisions. In contrast, polymorphisms in DNA repair genes directly determine repair capacity and thus chemotherapy responsiveness. Furthermore, the effects of germline SNPs may be masked in core effector genes such as TP53 by dominant somatic mutations. Given the high replication stress and genotoxic therapy exposure in AML, module-specific dependencies within the DDR network may account for the heterogeneous clinical associations observed.
The MRN ternary complex, composed of MRE11, RAD50 and NBS1, is one of the most critical damage sensors in the DDR pathway, playing a central role in recognizing DNA double-strand breaks, initiating signal transduction, and maintaining genomic stability [16, 17]. Our study found that the TC/CC genotypes of the MRE11 rs2155209 polymorphism served as protective factors against hyperleukocytosis in AML patients. Furthermore, survival analysis revealed that carriers of the MRE11 rs2155209 variant alleles had a longer OS. Currently, research on MRE11 rs2155209 in hematologic malignancies is limited, and studies in other cancers, such as bladder or breast cancer, have not identified this SNP as a protective factor against malignancy [18, 19]. Functionally, rs2155209, located in the non-coding region of the MRE11 gene, may upregulate MRE11 expression by influencing mRNA splicing or translational efficiency, thereby promoting apoptosis of AML cells in response to chemotherapeutic agents. Conversely, the CG/GG genotypes of the NBS1 rs1805794 polymorphism were identified as risk factors for hyperleukocytosis. Previous studies have reported inconsistent associations between NBS1 rs1805794 and susceptibility to hematologic malignancies, potentially illustrating the functional heterogeneity of this locus across different tumor types and genetic backgrounds [20, 21]. Rs1805794 is located in the BRCT domain of NBS1; the substitution of Gln for Glu may weaken the binding affinity between NBS1 and MRE11, thereby reducing the efficiency of DNA damage signal transduction.
The CHEK1, CHEK2, RAD17 and BRCA1 genes play key roles in the signal transduction and amplification module of the DDR pathway. Our study identified multiple significant associations within this module. For CHEK1 rs521102, the AA genotype served as a risk factor for thrombocytopenia but was associated with prolonged OS, suggesting a complex role where this variant may influence platelet production while simultaneously enhancing chemotherapy sensitivity [22, 23]. The CHEK2 rs738722 TT genotype has been identified as a risk factor for hyperleukocytosis, consistent with findings from multiple studies suggesting its potential association with poor cancer prognosis [24]. This effect may occur through modulation of CHEK2 activation efficiency, thereby impacting downstream p53-dependent apoptotic pathways. For RAD17 rs1045051, the GT/GG genotype emerged as a risk factor for AML relapse, which might be explained by impaired checkpoint function that reduces the effective elimination of residual leukemic clones following chemotherapy. We also identified the BRCA1 rs1799949 GA/AA genotype as a protective factor for high BM blast percentage. BRCA1 is a multifaceted tumor suppressor involved in both DNA damage signaling and homologous recombination repair [25, 26]. Although rs1799949 is a synonymous variant located in exon 11, it may up-regulate BRCA1 expression through effects on mRNA stability or splicing efficiency, thereby modulating the efficiency of both checkpoint activation and downstream repair processes [27]. Functionally, this variant could enhance the DNA damage response capacity of leukemic cells, preventing against more aggressive disease features.
In contrast, SNPs in the core effector genes TP53 and CDKN1A showed no significant associations in our study, which may be caused by several factors. Somatic TP53 mutations occur at high frequency in AML, particularly in cases with complex karyotypes [28, 29]. The dominant-negative or loss-of-function effects of TP53 mutations could easily overshadow any subtle influence of germline variants. Besides, the DDR signaling cascade contains considerable functional redundancy [30]. The ATM/CHK2/p53 axis operates in parallel with other DNA damage checkpoints, and p21, encoded by CDKN1A, can be regulated through p53-independent and post-transcriptional or post-translational mechanisms, providing compensatory routes that buffer the impact of germline SNPs in single effector genes [31, 32]. Additionally, because these pivotal tumor-suppressor genes are under strong purifying selection, common germline variants are likely to be functionally mild, further reducing their detectability in association studies of the present sample size [33].
The DNA repair module serves as the final executor of the DDR pathway, directly determining whether damaged cells undergo apoptosis or survive with genomic alterations [34]. Our study identified significant associations of AML course and 2 key repair genes, LIG3 and MGMT. For LIG3 rs3744356, the TT genotypes were associated with multiple adverse clinical features, including elevated WBC counts, lower CR rate following induction chemotherapy, and shorter OS. LIG3 plays a critical role in base excision repair and alternative non-homologous end joining; functionally, rs3744356 may upregulate LIG3 expression by influencing mRNA splicing or stability, thereby enhancing the repair capacity of leukemic cells against chemotherapy-induced DNA damage and promoting drug resistance [35]. For MGMT rs2296675, the variant allele was identified as a risk factor for shorter OS. MGMT directly repairs alkylating agent-induced DNA damage, and rs2296675, though a synonymous variant, may influence MGMT expression through codon usage bias or mRNA stability, potentially allowing damaged cells to survive and proliferate [36], ultimately leading to a poorer prognosis in AML. Currently, studies on both LIG3 rs3744356 and MGMT rs2296675 in AML are limited; our findings provide the evidence linking these variants to clinical outcomes in AML. Together with our observations in the damage sensing and signal transduction modules, these results support the concept that genetic variation across the entire DDR cascade—from damage recognition to final repair—collectively shapes AML aggressiveness, treatment response, and patient survival.
In summary, this study systematically evaluated the association between a multi-gene SNP panel spanning the DDR pathway and AML aggressiveness and prognosis. Our findings highlight that germline variants in damage sensing, signal transduction, and DNA repair modules play significant roles in AML development and chemosensitivity, while the contribution of SNPs in effector genes such as TP53 and CDKN1A may be limited in this context. Given the widespread accessibility of SNP genotyping assays and the relatively high frequency of these variants in the Chinese population, prospective evaluation of these SNPs in clinical laboratories is entirely feasible. However, several limitations should be acknowledged. The modest sample size may limit the statistical power for multivariate analyses and subgroup assessments. The use of dbSNP database allele frequencies as controls, rather than a matched healthy cohort, may introduce population stratification bias. Additionally, the precise molecular mechanisms by which these SNPs influence DDR function remain to be elucidated, and somatic mutation data were not integrated into the current analysis. Future studies with larger, independent cohorts and well-matched controls are warranted to validate our findings. Functional experiments are also needed to clarify how these variants modulate DDR pathway activity and ultimately influence AML progression and treatment response. Despite these limitations, our results provide a foundation for developing a DDR-based polygenic risk score model and open new avenues for integrating germline genetic profiling into precision medicine strategies for AML.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (No. 82370173, No. 32241005).
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author contributions
Daoxin Ma conceptualized this study. Amin Zhang and Wancheng Liu acquired samples and analyzed data. Amin Zhang and Hanyang Wu performed the experiment and analyzed the data. Amin Zhang Hanyang Wu, Wancheng Liu, Daoxin Ma wrote and revised the manuscript. All authors studied and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Miles LA, Bowman RL, Merlinsky TR, Csete IS, Ooi AT, Durruthy-Durruthy R. et al. Single-cell mutation analysis of clonal evolution in myeloid malignancies. Nature. 2020;587:477-82
2. Song Z, Cui Y, Xin L, Xiao R, Feng J, Li C. et al. Mechano-oncogenic cytoskeletal remodeling drives leukemic transformation with mitochondrial vesicle-mediated STING activation. Cell Stem Cell. 2025;32:581-97 e11
3. McMahon CM, Perl AE. Management of primary refractory acute myeloid leukemia in the era of targeted therapies. Leuk Lymphoma. 2019;60:583-97
4. Bouchla A, Sotiropoulou CD, Esteb C, Loupis T, Papageorgiou SG, Deliconstantinos GG. et al. Silencing of the DNA damage repair regulator PPP1R15A sensitizes acute myeloid leukemia cells to chemotherapy. Ann Hematol. 2024;103:2853-63
5. Shastri VM, Chauhan L, Gbadamosi M, Alonzo TA, Wang YC, Aplenc R. et al. DNA Damage Response Pharmacogenomic (DDR_PGx) Score Predicts Response to Chemotherapy Consisting of Gemtuzumab Ozogamicin in Pediatric AML: A Report from the Children's Oncology Group. Clin Cancer Res. 2025;31:890-8
6. Kawano G, Ikeda R, Ishihara D, Shima T, Sakoda T, Yamamoto S. et al. Guanine nucleotides drive ribosome biogenesis and glycolytic reprogramming in acute myeloid leukemia stem cells. Blood. 2026;147:768-82
7. Vergez F, Largeaud L, Bertoli S, Nicolau ML, Rieu JB, Vergnolle I. et al. Phenotypically-defined stages of leukemia arrest predict main driver mutations subgroups, and outcome in acute myeloid leukemia. Blood Cancer J. 2022;12:117
8. Kishtagari A, Levine RL. The Role of Somatic Mutations in Acute Myeloid Leukemia Pathogenesis. Cold Spring Harb Perspect Med. 2021 11
9. Cuartero S, Innes AJ, Merkenschlager M. Towards a Better Understanding of Cohesin Mutations in AML. Front Oncol. 2019;9:867
10. Shimony S, Stahl M, Stone RM. Acute myeloid leukemia: 2023 update on diagnosis, risk-stratification, and management. Am J Hematol. 2023;98:502-26
11. Warner E, Herberts C, Fu S, Yip S, Wong A, Wang G. et al. BRCA2, ATM, and CDK12 Defects Differentially Shape Prostate Tumor Driver Genomics and Clinical Aggression. Clin Cancer Res. 2021;27:1650-62
12. Roy R, Chun J, Powell SN. BRCA1 and BRCA2: different roles in a common pathway of genome protection. Nat Rev Cancer. 2011;12:68-78
13. Hall T, Gurbuxani S, Crispino JD. Malignant progression of preleukemic disorders. Blood. 2024;143:2245-55
14. Arends CM, Kopp K, Hablesreiter R, Estrada N, Christen F, Moll UM. et al. Dynamics of clonal hematopoiesis under DNA-damaging treatment in patients with ovarian cancer. Leukemia. 2024;38:1378-89
15. Ellis NA, Huo D, Yildiz O, Worrillow LJ, Banerjee M, Le Beau MM. et al. MDM2 SNP309 and TP53 Arg72Pro interact to alter therapy-related acute myeloid leukemia susceptibility. Blood. 2008;112:741-9
16. Bian L, Meng Y, Zhang M, Li D. MRE11-RAD50-NBS1 complex alterations and DNA damage response: implications for cancer treatment. Mol Cancer. 2019;18:169
17. Situ Y, Chung L, Lee CS, Ho V. MRN (MRE11-RAD50-NBS1) Complex in Human Cancer and Prognostic Implications in Colorectal Cancer. Int J Mol Sci. 2019 20
18. Wu Z, Wang P, Song C, Wang K, Yan R, Li J. et al. Evaluation of miRNA-binding-site SNPs of MRE11A, NBS1, RAD51 and RAD52 involved in HRR pathway genes and risk of breast cancer in China. Mol Genet Genomics. 2015;290:1141-53
19. Liao CH, Chang WS, Shih HY, Wang YC, Hsu CL, Chang SY. et al. Contribution of MRE11, RAD50, and NBS1 Genotypes to Bladder Cancer Susceptibility. Cancer Genomics Proteomics. 2025;22:575-91
20. Jiang L, Liang J, Jiang M, Yu X, Zheng J, Liu H. et al. Functional polymorphisms in the NBS1 gene and acute lymphoblastic leukemia susceptibility in a Chinese population. Eur J Haematol. 2011;86:199-205
21. Li N, Xu Y, Zheng J, Jiang L, You Y, Wu H. et al. NBS1 rs1805794G>C polymorphism is associated with decreased risk of acute myeloid leukemia in a Chinese population. Mol Biol Rep. 2013;40:3749-56
22. David L, Fernandez-Vidal A, Bertoli S, Grgurevic S, Lepage B, Deshaies D. et al. CHK1 as a therapeutic target to bypass chemoresistance in AML. Sci Signal. 2016;9:ra90
23. Qiu Z, Oleinick NL, Zhang J. ATR/CHK1 inhibitors and cancer therapy. Radiother Oncol. 2018;126:450-64
24. Tu R, Zhong D, Li P, Li Y, Chen Z, Hu F. et al. PTPN13 rs989902 and CHEK2 rs738722 are associated with esophageal cancer. Ann Med. 2023;55:2281659
25. Zhao W, Wiese C, Kwon Y, Hromas R, Sung P. The BRCA Tumor Suppressor Network in Chromosome Damage Repair by Homologous Recombination. Annu Rev Biochem. 2019;88:221-45
26. Russi M, Marson D, Fermeglia A, Aulic S, Fermeglia M, Laurini E. et al. The fellowship of the RING: BRCA1, its partner BARD1 and their liaison in DNA repair and cancer. Pharmacol Ther. 2022;232:108009
27. Chen X, Truong TT, Weaver J, Bove BA, Cattie K, Armstrong BA. et al. Intronic alterations in BRCA1 and BRCA2: effect on mRNA splicing fidelity and expression. Hum Mutat. 2006;27:427-35
28. Fedenko A, Czapinska H, Krämer A, Stölzel F, Bochtler T, Bochtler M. Etiology of TP53 mutated complex karyotype acute myeloid leukemia. Leukemia. 2026;40:444-8
29. Lodé L, Ameur A, Coste T, Ménard A, Richebourg S, Gaillard JB. et al. Single-molecule DNA sequencing of acute myeloid leukemia and myelodysplastic syndromes with multiple TP53 alterations. Haematologica. 2018;103:e13-e6
30. Smith HL, Southgate H, Tweddle DA, Curtin NJ. DNA damage checkpoint kinases in cancer. Expert reviews in molecular medicine. 2020;22:e2
31. de Queiroz RM, Moon SH, Prives C. O-GlcNAc transferase regulates p21 protein levels and cell proliferation through the FoxM1-Skp2 axis in a p53-independent manner. The Journal of biological chemistry. 2022;298:102289
32. Jung YS, Qian Y, Chen X. Examination of the expanding pathways for the regulation of p21 expression and activity. Cellular signalling. 2010;22:1003-12
33. Khan MM, Rydén AM, Chowdhury MS, Hasan MA, Kazi JU. Maximum likelihood analysis of mammalian p53 indicates the presence of positively selected sites and higher tumorigenic mutations in purifying sites. Gene. 2011;483:29-35
34. Chiolo I, Altmeyer M, Legube G, Mekhail K. Nuclear and genome dynamics underlying DNA double-strand break repair. Nat Rev Mol Cell Biol. 2025;26:538-57
35. Hua RX, Zhuo Z, Zhu J, Zhang SD, Xue WQ, Li XZ. et al. LIG3 gene polymorphisms and risk of gastric cancer in a Southern Chinese population. Gene. 2019;705:90-4
36. Kaina B, Christmann M, Naumann S, Roos WP. MGMT: key node in the battle against genotoxicity, carcinogenicity and apoptosis induced by alkylating agents. DNA Repair (Amst). 2007;6:1079-99
Author contact
![]() Corresponding authors: Daoxin Ma, Professor (Email: daoxinmaedu.cn), Wancheng Liu, Professor (Email: LWC15190com).
Corresponding authors: Daoxin Ma, Professor (Email: daoxinmaedu.cn), Wancheng Liu, Professor (Email: LWC15190com).